
回答这个问题前,我们需要弄清楚:人类的寿命为什么会有限?
20世纪60年代以前,人们一直相信脊椎动物的细胞拥有无限分裂的能力。直到1961年,美国微生物学家列奥那多·海弗里克的发现,打破了人们的幻想。
通过实验观察,海弗里克发现,正常细胞在体外的分裂次数存在一个40~60次的上限。这个上限,又被称为海弗里克极限[1]。
20世纪70年代,科学家伊丽莎白·布莱克本(Elizabeth Blackburn)等人发现了位于染色体末端的端粒结构。
人类端粒
端粒可以保护染色体,并保证基因复制时的稳定性,但端粒在每次复制的时候会丢失一部分。当丢失的端粒达到极限,细胞也就不再分裂[2]。
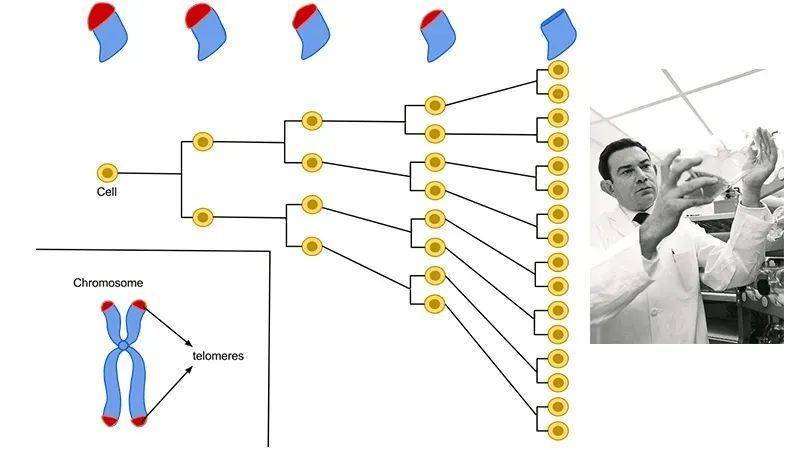
端粒的发现,完美地解释了海弗里克极限。
伊丽莎白布莱克本等人,还发现了能修复端粒的端粒酶。
端粒酶分子结构
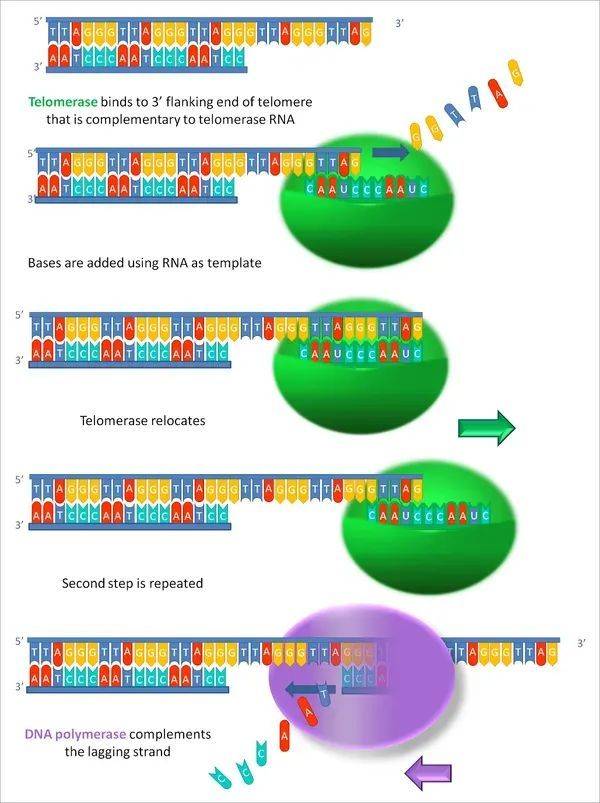
端粒酶和DNA聚合酶制造端粒的顺序
不同细胞海弗里克极限的长短,受限于细胞端粒的长度以及修复能力。胰岛细胞的海弗里克极限只有10余次,而造血干细胞的海弗里克极限可达到100多次。
进入21世纪之后,端粒的研究已经成为了生物学的热门领域。2009年,伊丽莎白布莱克本等人被授予诺贝尔生理学或医学奖[3]。
利用端粒酶修复端粒,是否就能打破海弗里克极限呢?
胚胎干细胞和生殖细胞之所以能无限分裂,正在于端粒酶对端粒的修复。但在正常细胞中,如果端粒酶过于活跃,则会导致癌症。而癌细胞之所以能无限分裂,同样在于端粒酶对端粒的无限修复。
可否有可能突破海弗里克极限,又不提高癌症的发生呢?
这是可以的,2000年前后,研究者引入外源端粒酶对细胞染色体端粒进行修复(异位表达)[4] [5],打造出了没有癌化的永生细胞系。端粒酶的异位表达拥有广阔的
在遥远的将来,利用端粒酶的异位表达来实现永生,并非没有可能性。
在无脊椎动物中,不少动物都拥有无限的理论寿命,例如水螅、龙虾等。其中,龙虾就有无限修复端粒的能力。
然而限制永生的,可不仅仅只有端粒。
2013年,一篇名为《The hallmarks of aging(衰老的特征)》的文章刊登在了《Cell》杂志上,对人类衰老研究进行了一系列的经典总结[6]。
该文列举了影响人体衰老的九大特征,除了端粒损耗外,还有基因组不稳定性、表观遗传学改变、蛋白内稳态失衡、营养感应失调、线粒体功能异常、细胞衰老、干细胞耗竭,以及细胞间通讯改变。
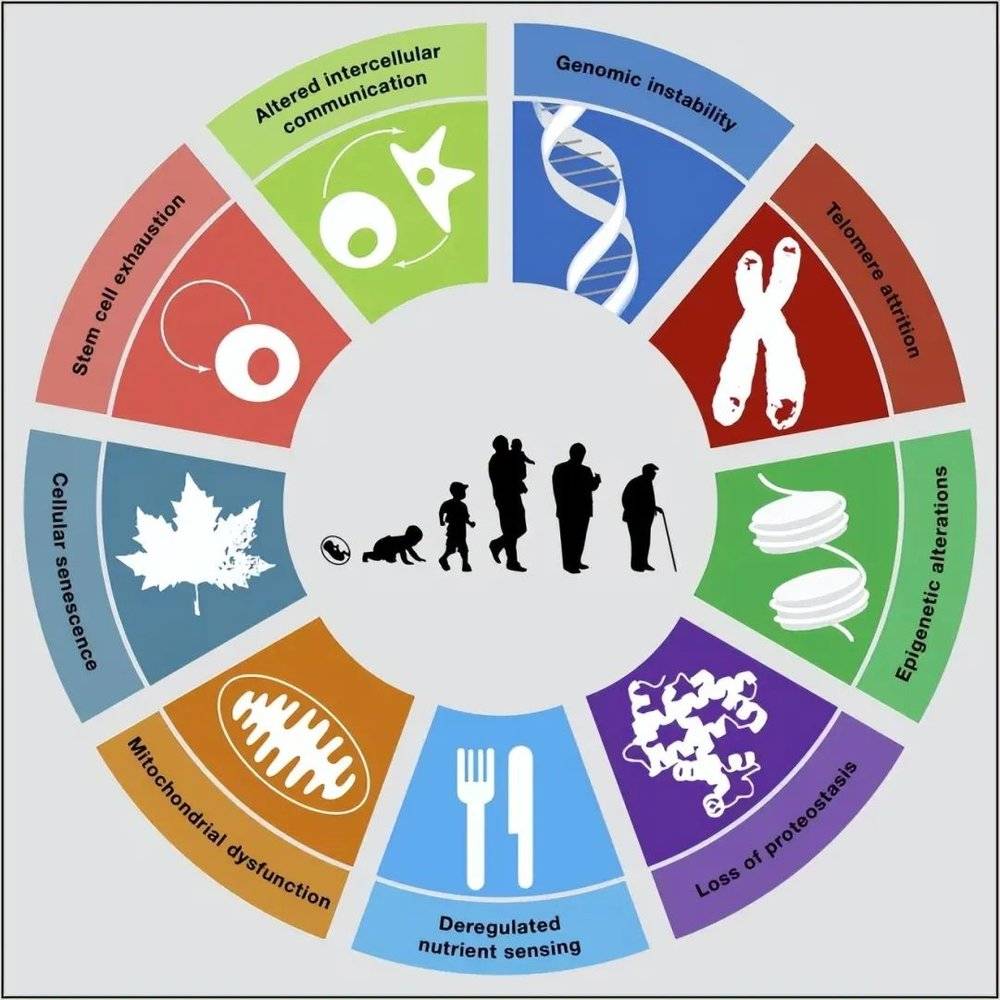
九种因素互相影响,共同决定了人体的衰老,但最终起到决定性作用的还是在基因层面。
从根本上来说,其实这都是人生数十年来,从细胞、分子的微观层面,破坏/损伤的不可逆积累。
人类细胞达到100万亿,每天都会更新3300亿个,要从细胞、分子层面上修复,且不说难度,单单从总工作量的角度上说,都是一个天文数字。
永生技术的根本性难题,犹如天堑一般挡在了人类的身前。
也正是在这个大前提下,很多人把数字永生视为无法实现生物永生的替代选择。
数字永生真的能够实现,作为生物永生的替代选择吗?
真正意义上数字永生(而不仅仅是电子数据备份)的前提,是能做到真正的意识上传。
然而在意识上传之前,却必须面对一个现今无法完全解答的问题:
意识究竟是什么?
高阶理论(HOTs)、全局工作空间理论(GWT)、信息整合理论(IIT)、再入和预测处理理论,是解释意识的四大主要理论,然而令人类尴尬的是,这些理论目前来说,都无法真正的验证。
甚至因为相信的人太多,信息整合理论(IIT)还被124个科学家“盲目”打上了“伪科学”的标签[7]。
但其实,我们也可以不用管意识理论之争。至少神经科学发展100年以来,存在一些基本的共识:
一般把意识看作大脑的一种物质性、生理性功能。
虽然我们并没有完全弄清楚意识究竟是什么,但有一些实验至少证明了,意识无法脱离大脑的物质基础。
早在20世纪80年代,心理学家本杰明•李贝特便做了一个著名的实验[8] [9]。
他们让5个左撇子的大学生坐在躺椅上,并告诉他们用1~2秒的时间放松头部、颈部,以及前臂肌肉。但在决定做这一件事情之前,他们需要突然快速动一根手指或手腕。当他们活动手指的时候,不要有任何的预先计划或刻意关注,随机重复40次。
在这些大学生进行这些动作时,研究人员测量了三个变量:
1. 贴在前臂的电极,记录手指动作开始的时间。
2. 贴在头皮上的电极,测试动作开始时大脑的预备电位。
3. 感受到行为冲动(想动手指)时,喊出屏幕中钟表的“时间”,从而测出决定时刻。
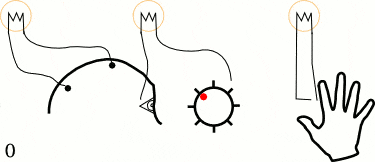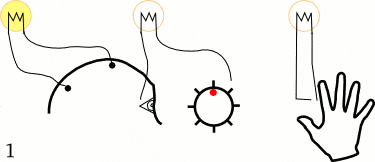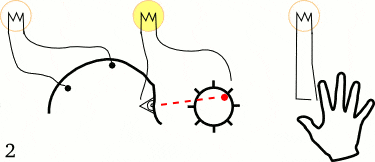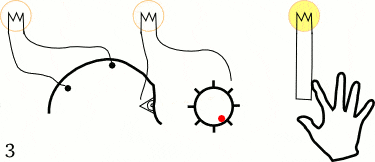
经过多达几百次实验,李贝特最终发现,决定时刻出现在大脑预备电位之后,平均时间间隔为350ms。
也就是说,当我们决定做某一件事情之前的1/3秒左右,大脑就已经发起动作了。
因此,有研究者悲观地认为人类并没有自由意识。
不过在进一步的实验中,李贝特让被试大学生,在做决定之后否决行为。
虽然大脑出现了预备动作电位,但最终阻止了动作,并且没有检测到手上的电位。近年来,也有研究进一步表明,当大脑动作电位出现后的一定时间内,可以进行否决,但距离动作时间足够近时,否决的成功率便会大大降低[10]。
这说明,无论人类有没有自由意识,但在一定时间内都有否决的自由。
40年来,有大量的研究支持李贝特实验中意识决定晚于大脑动作的结果。延后时间短则数百毫秒,最长甚至可达10余秒[11] [12] [13]。
在一项实验中,经颅磁刺激改变受试者的左右手使用习惯后,受试者依旧认为自己的选择经过了自由意识的决定[14]。在某些实验设计下,当出现无意识判断或冲动行为,受试者也会认为是自己的决策行为[15]。
直接对高级皮层进行刺激,受试者则可能出现错误的意识判断。例如,他们可能认为自己做出了某种动作,但实际并非发生[16]。
这似乎更加肯定了人类没有自由意识。
然而,最近10多年一些研究者却有了进一步的发现,有研究者否定了人类没有自由意志的看法。
2009年,有人把李贝特的经典实验,修改为播放一段音频,然后让志愿者决定是否敲击一个键。研究发现,不管志愿者是否真的选择了敲击,两种情况下都有相同的大脑预备电位。
这表明,大脑预备电位并不表明已经做出了决定。
当然,志愿者即刻决定是用左手还是右手按键时,大脑的早期动作电位同样没有什么区别,这说明大脑早期产生的动作电位,可能是注意到信号或者对信息的预处理。
近年来,越来越多更精确的方法证明了意识决定不是瞬间出现的,而是逐步建立起来的。因此有研究者认为,决策结果的早期神经标记不是无意识的,而是简单地反映了有意识的目标评估阶段,这些阶段还不是最终的,在达成最终意识活动之前,这个决策可以终止或改变[17]。
总之,一个动作可能在我们的“意识”意识到它之前就已经开始了,并不意味着我们的意识不能批准、修改或者取消这个动作。
以上这些实验,使我们可以在不谈具体意识理论的前提下,进行这样的判断:
意识是大脑神经活动之后的产物,它并没有先决性,也不能单独存在。并不存在一段单独的信息(甚至量子态的意识/灵魂),用来进行意识上传。
要做到真正的意识上传,本质上是需要维持意识的连续性。
可喜的是,意识的连续性,可以通过物质的连续性来维持。
我们大脑这种物质,在我们成长过程中,本身就是不断连续变化,微观结构不断持续替换的过程。我们的自传体记忆,维持着我们意识和自我意识的连续变化,让我们有意识体验和自我体验。
而这些连续变化的信息,正是成千上万的电化学过程,是可以通过电子硬件来进行复制或替换的。
也就是说,要做到真正的意识上传,那么就需要打造出真正可替的电子神经元,然后如同一个人的成长过程那样,逐步替换掉神经元,维持自体记忆的连续性,这样才能真正地做到意识上传。
也就是说真正意义上的数字永生,需要的是细胞层面上的等效替换,这就需要人类分子纳米机器人技术已经发展到极高的地步。
前面提到,在生物水平上,人类之所以不能永生,前文提到的9个原因(基因组不稳定性、表观遗传学改变、蛋白内稳态失衡、营养感应失调、线粒体功能异常、细胞衰老、干细胞耗竭),本质上都体现在一种破坏的积累。
如此高水平的纳米机器人技术,已经完全能够做到在分子层面随时随地地修复破坏。
不仅能够让人永生,甚至能够真正做到永葆青春,使得生理年龄维持恒定不变。
也就是说,很有可能,真正的数字永生是以生物永生已经实现为前提的。
它自然也就失去了生物永生走不通时的替代意义。而像电影《流浪地球2》中的数字人,我更宁愿称之为电子复制人,而非真正意义上的数字永生。
当你实现生物永生,你的身体内,随时随地都存在着大量的可控纳米机器人。
人体细胞达到100万亿,哪怕1%都是1万亿的纳米机器人。
所有纳米机器人的精准控制,至少需要一台超算来维持,需要现今尚不存在的匹配人体的优秀人工智能算法,考虑到人体本身是持续维持平衡的耗散系统,生化内环境的瞬息万变,这些纳米机器人的控制中心,也需要持续地进行深度学习。
对任何一个人永生的持续维持,都是以极高的成本为代价。
而为了更好地匹配机器,人体改造也是在所难免。再加上机器的维护成本有可能比有机体更低,未来也可能会有更多的人选择用机器替代身体。
随着时代的发展,量变到质变。生物永生和数字永生的界限,有可能会变得越来越模糊,最终殊途同归。
在这种情况下,生物体的形态,甚至可以做到自由设计的地步。
那么,在最后我们是否可以提出这样的一个假想:无论生命/文明以何种方式起源,宇宙中的终极生命形态,在物质上的差异都会极小。最大的差别,可能只是文化差异,对生命外观形象的个体/群体性偏好。
以上超出现有理论的部分,皆是个人观点。
参考文献:
[1] Shay J W, Wright W E. Hayflick, his limit, and cellular ageing[J]. Nature reviews Molecular cell biology, 2000, 1(1): 72-76.
[2] Blackburn E H. Switching and signaling at the telomere[J]. Cell, 2001, 106(6): 661-673.
[3] https://www.nobelprize.org/prizes/medicine/2009/summary/
[4] Counter C M, Meyerson M, Eaton E N, et al. Telomerase activity is restored in human cells by ectopic expression of hTERT (hEST2), the catalytic subunit of telomerase[J]. Oncogene, 1998, 16(9): 1217-1222.
[5] Hooijberg E, Ruizendaal J J, Snijders P J F, et al. Immortalization of human CD8+ T cell clones by ectopic expression of telomerase reverse transcriptase[J]. The Journal of Immunology, 2000, 165(8): 4239-4245.
[6] López-Otín C, Blasco M A, Partridge L, et al. The hallmarks of aging[J]. Cell, 2013, 153(6): 1194-1217.
[7] Fleming S, Frith C, Goodale M, et al. The Integrated Information Theory of Consciousness as Pseudoscience[J]. 2023.
[8] Libet B , Gleason C A , Wright E W , et al. TIME OF CONSCIOUS INTENTION TO ACT IN RELATION TO ONSET OF CEREBRAL ACTIVITY (READINESS-POTENTIAL)[J]. Brain, 1983, 106(3):623-642.
[9] Libet B . Unconscious cerebral initiative and the role of conscious will in voluntary action[J]. Behavioral & Brain ences, 1985, 8(4):529-539.
[10] Schultze-Kraft M , D Birman, Rusconi M , et al. The point of no return in vetoing self-initiated movements[J]. Proc Natl Acad Sci U S A, 2016, 113(4):1080-1085.
[11] Matsuhashi M , Hallett M . The timing of the conscious intention to move[J]. European Journal of Neuroence, 2010, 28(11):2344-2351.
[12] Soon, C., Brass, M., Heinze, HJ. et al. Unconscious determinants of free decisions in the human brain. Nat Neurosci 11, 543–545 (2008).
[13] Soon C S , He A H , Bode S , et al. Predicting free choices for abstract intentions[J]. Proceedings of the National Academy of Sciences, 2013, 110(15).
[14] Brasil-Neto J P , Pascual-Leone A , Valls-Sole J , et al. Focal transcranial magnetic stimulation and response bias in a forced-choice task.[J]. J Neurol Neurosurg Psychiatry, 1992, 55(10):964-966.
[15] S Kühn, Brass M . Retrospective construction of the judgement of free choice[J]. Consciousness & Cognition, 2009, 18(1):12-21.
[16] Desmurget, M.; Reilly, K. T.; Richard, N.; Szathmari, A.; Mottolese, C.; Sirigu, A. (2009). "Movement Intention After Parietal Cortex Stimulation in Humans". Science. 324 (5928): 811–813.
[17] Guggisberg A G , Ana?S M . Timing and Awareness of Movement Decisions: Does Consciousness Really Come Too Late?[J]. Frontiers in Human Neuroscience, 2013, 7.
本文来自微信公众号:瞻云(ID:zhanyun2028),作者:瞻云