
本文来自微信公众号:神经现实(ID:neureality),作者:杞人,编辑:阿莫東森,原文标题:《酒神,你可是理智的囚徒?》,头图来自:《比利·林恩的中场战事》
如果非要问什么是情感的话,它应该包括我们的内在感受、生理状态和表达情感的行为。除去每日的喜怒哀乐、嗔痴怒骂,生理上的疼痛也能诱导出情感。迄今为止,我们的对情感的实验告诉我们,这些情感体验将生活中每一件大大小小的事都分类为好的,坏的,中性的,从而引发或者调节我们应对这些事件的行为。除此之外,情感还控制着我们如何感受这个世界,影响着我们的记忆和学习。
从古至今,无论是西方文化,还是中国文化,理智一直都和感性对立。有人曾推崇绝对的理性,扼杀内心的冲动,也有人推崇绝对的感性,让一切荒谬都臣服于个人的情感。可是,随着科学技术的发展,最近的研究发现情感很难与理性分开。它们紧密相连,互为因果。
找一找,情感脑区在哪里?
科学家一直在寻找负责情感的脑区,可惜多年寻觅终是难以给出一个确切的答案。在1878年,法国神经学家保罗·布罗卡(Paul Broca)描述了一组在大脑内侧和周围的皮质都不太一样的皮质区,并将其命名为边缘叶(limbic lobe)。当时他并没有意识到,他所定义的区域中有很多脑区都是和情感紧密相关的。
在20世纪30年代, 美国神经学家詹姆士·帕佩兹(James Papez)描述了帕佩兹回路(Papez circuit),其中有很多脑区都属于布罗卡描述的边缘叶的一部分。其中下丘脑(hypothalamus)控制着表达情感的行为,而新皮层(neocortex)和下丘脑的连接允许情感的经历和情感的表达相互控制。
然而后来的研究发现,帕佩兹回路少了很多重要的控制情感的脑区,例如杏仁核(amygdala)。随着更多脑区的加入,人们沿用了布罗卡的命名,并对这些控制情感的脑区起了一个统称:边缘系统(limbic system)。
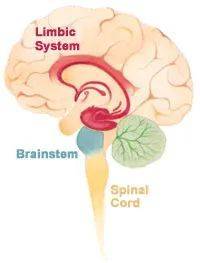
可是事情真的这么简单吗?我们可以用一个系统来描述我们的情感么?越来越多的证据显示,就像记忆一样,情感是一个全脑参与的过程,包括了认知和感官知觉。“边缘系统”没有办法解释情感中的所有元素。
意识到了没有一个固定的“情感脑区”,研究人员转而询问当人类在有不同情感的时候,会不会有一个普遍的全脑激活规律?传统的功能性核磁共振(fMRI)通过测量脑部血液和氧气分布来对比哪些脑区在进行一项任务的时候更加活跃,更加活跃的脑区会有更多的血流量。这个技术的空间单位是体素(voxel),一个体素大概可以包含63万个神经元。单变量fMRI(也就是我们经常用到的fMRI)同时分析一个脑区的几千个体素,因此细节的对比在这种方法里变得十分困难。
随着技术的发展,多体素模式分析(multivoxel pattern analysis/MVPA)可以测量每一个体素单独的血液和氧气分布,所以在情感网络的学习上,这项技术对寻找一个普遍的网络激活十分有帮助。另一种多体素模式分析利用相关矩阵来对比在同一认知任务中,多脑区激活规律的相似和不同,所以它可以很好的找到刺激和相对应的大脑反应的关系。这种技术叫做代表性相似分析(RSA,representational similarity analysis)。
在2013年,卡萨姆等人(Kassam et al., 2013)利用多体素模式分析来研究情感。通过机器学习,分类器先学会了在不同情感状态下,被试的脑区激活是有怎样的一个规律。在这个基础上,分类器可以利用新的激活规律准确地预测同一个人此时的情感是什么、这种情感有多强烈。除此之外,它们还可以相对准确地利用现有的知识预测另一个人的情感。
值得一提的是,在研究不同人在同样的情感刺激下有哪些共同的脑区激活时,他们发现有很多区域不在传统的边缘系统里。类似的研究还有很多,他们都指向同一个结论:人类情感导致的脑区激活存在可以预测的规律,并且这些被激活的脑区远远超过传统的边缘系统,包括了负责认知的脑区。
冲动,从世界接触你皮肤的一刻便开始
如果你的身体是一个房子,那么房子上面的窗户就是你的各种感官,它们片面地提取世界中的各类信息,然后情感将这些信息简单地分成好的或坏的。
例如疼痛,人类对疼痛的感知并不是由一个专门处理疼痛的脑区决定的,相反,这种感知从受伤感受器(nociceptor)被激活的一瞬间就注定产生疼痛的知觉。伤害感受器向中枢神经发送信号,其中被髓鞘包裹着的快速神经(myelinated fast fibre)告诉大脑疼痛区域的位置,而没有被髓鞘包裹的慢速神经(unmyelinated slow fibre)传递一种更加扩散的疼痛之感。有没有磕过脚趾?那你一定知道疼痛先是尖锐而可以忍受的,可片刻过后,那种疼痛就仿佛不能忍受,它使你缓缓蹲下,表情狰狞。
在步入21世纪以后,研究者发现了一个与疼痛很相像的知觉:“令人愉快的触碰”(Olausson et al., 2002; Loken et al., 2009)。如果将小刷子用一个被被试评为“最舒适”的速度轻轻刷过他/她的皮肤,C类触觉传入细胞(C-tactile afferents)发射动作电位的速率是最高的。这些研究体现了令人愉快的体感在社会动物中的重要性。
上面所讨论的两个系统都将信息传递给眶额叶(OFC,orbitofrontal cortex),一个研究者们长久以来认为和情感经历相关的脑区。所以虽然情感经历并不是在皮肤上就产生了,但是产生什么样的感情在皮肤上就已经被决定,而不是等到信息达到了大脑才被决定。
味觉系统就像触觉系统一样,在我们探测化学物质的时刻,情感的好坏便已经被决定。在前腹脑岛(anteroventral insula)和后侧眶额叶(posterior OFC)被认为是有可能处理味觉的第一和第二皮质,而令人愉快的和令人厌恶的味觉刺激在这两个区域里拥有不同的激活规律(Small et al., 2003)。最恶心的味道和最令人愉快的味道的激活规律最为不同,而激活规律的相似度随着情感的相似程度的增加而增加。
与以上系统不同,视觉系统无法在视网膜的水平上就将令人愉快的和不愉快的信息区分开来(强光照射产生疼痛的过程可以算作疼痛的感知,从另一个角度来讲可以和视觉区分开来)。这些信息经过整合,在更高阶的视觉皮质(higher-order visual cortex)形成了物体的特征,此时这些特征是好的还是坏的才会被区分。
感官知觉在情感体验中担任很重要的角色,因为人体的感官系统从一开始就将会产生积极情感和消极情感的刺激分别开来,从而允许大脑做出不同的回应。它展示了我们的情感体验不是机械化的“这里有一个东西,我感觉很开心”,而是一种更加流畅的体验:“这里有一个让我感觉很开心的东西”。
在大多数情况下,一个刺激的好坏标准是由基因决定的,因此后天的环境对标准影响并不会很大,而这种统一的衡量标准允许我们在没有太多思考的情况下,对一切不好的刺激快速地做出反应,避免伤害进一步发生。无意识情感(unconscious emotion)提供了很好的一个例子:即使被试不承认看到过一个快速闪过的愤怒表情,他们的手心依然会出汗。
我心有千层浪,许是野马被困住太久
不可否认,情感在很大程度上控制我们的行为。如果你去看那些经典的控制情感的脑区(例如杏仁核),你会发现它们和控制人体激素分泌,自主神经系统(autonomic nervous system)的下丘脑,控制动机的脑区都有很紧密的连接。因此情感可以控制我们的生理状态,从而帮助我们更好地应对外界的威胁。
更重要的是,情感还会转移我们的注意力,影响记忆的形成和巩固,所以我们会记得那些不好的事情,并且把注意力快速的放在与这些记忆有关的刺激上,避免它再一次发生。对杏仁核的研究虽然将它经常和恐惧、侵略性紧紧联系在一起,但是在不同的情感中它多少都有参与。
在恐惧中,杏仁核接收来自海马体(hippocampus),新皮质(neocortex)和扣带回(cingulate gyrus)的有关恐惧的信息,并将信息传递给下丘脑和导水管周围灰质(periaqueductal grey matter/PAG)。下丘脑通过刺激垂体前叶(anterior pituitary)分泌激素并且激活交感神经系统(sympathetic nervous system)使心跳加快,心输出量增加,并分泌肾上腺素等等。而周围导水管灰质控制我们有意识的行为。
这一切都让我们更好地做好“战斗或逃跑”的反应(fight-or-flight response)。在积极的情绪当中,杏仁核和分泌多巴胺的伏隔核(nucleus accumben)的连接意味着情绪还调控我们的动机和行为。
我们的注意力也时时刻刻被情感影响着。杏仁核通过调节眼动,让人们把视线放在那些引起不好情绪的事物上,所以那些引起痛苦记忆的刺激会得到更多的注意,从而让人避免悲剧再一次发生。同理,如果人们能把注意力放在和奖励相关的刺激上,他们更有可能获得奖励。这种习惯往往会伴随人们很长时间,而且人生经历对这些刺激的调节起到着很大的作用。
这种功能如果运作不当便会导致一些问题,例如创伤后应激障碍(PTSD)和抑郁症(depression)。在创伤后应激障碍患者中,这种功能往往过于活跃。对于那些在阿富汗服过役并患有PTSD的战士来说,他们对战斗相关的刺激和没有打过仗的普通士兵相比,会有更多的视觉皮质的激活(Todd et al. 2015)。这意味着他们对战争相关的刺激会更加敏感,所以应激反应也会更大。在他们眼里,这个世界充满了危险。
在生活中,你会发现回想上星期早上开车时发生了什么十分困难,可是对于那些曾经经历了严重车祸的人来说,他们可以很清晰地回忆起车祸前发生了什么,即使很多年过去他们也都不会忘记。这是因为,我们的情感还会影响记忆的形成,相比于没有情感参与的事件,人们会倾向于更好地记住那些有感情的参与的事件。
这不仅是因为注意力被转移到那个引起情感的刺激上,情感的强烈程度对记忆的巩固也很重要。换言之,被情感控制的注意力帮助我们形成与情感刺激相关的记忆,而情感的强烈程度影响这个事件记忆的深刻程度。
虽然这种记忆往往和消极的情绪相关,积极的情绪也可以影响我们的记忆,尤其是联结式学习(associative learning)。人们可以更好地将快乐和其相关的事物结合起来,从而促进寻求奖励的行为。
杏仁核在这一过程中不出所料地继续担任着很重要的位置。海马体和杏仁核的连接允许人们更好的记住并提取和情感相关的经历,同时杏仁核还促进了条件增强(conditioned reinforcement),使人们可以更好地学习与条件刺激有关的知识,而伏隔核通过调节动机辅助杏仁核更好地完成这项任务。如果基底外侧杏仁核(basolateral amygdala)被损坏,巴甫洛夫式间接性恐惧条件反射(Pavlovian fear conditioning)就很难被形成,更不必提其相关的学习。
2014年,米斯科维奇和凯尔的实验支持了情感促进条件增强的理论(Miskovic and Keil, 2014)。如果被试看到轻微视觉提示的时候听到一声很大的令人不适的噪音,被试捕捉这些轻微的视觉提示的视觉灵敏度会增加,同时视觉皮层里也会有更大的激活。
显而易见,情感对于人们生理状态和行为的调控极其重要。我们认为理性和感性是对立的,这也许是因为人们追求情感冲动的时候往往会做出令人后悔的决定,故而情感被附上了“有缺陷”的标签。可是他们本质上更像是相辅相成、不分等级的,因为我们对世界的认知,我们的注意力,我们的记忆都被情感调控着,而当我们做出理性的决定时,很大程度上我们又会依赖于过去的经验和学习。
如果说情感是“有缺陷”的,那不如说理智的根本也是有缺陷的。可这并不意味着我们是情感的奴隶,人类在很多情况下会明知自己的决定将带来痛苦却依然选择去执行。这是因为我们可以对未来做出预测,而且我们追求的不光是短期的快乐,而是长期的积极的结果。
理智和情感的界限像生物里很多其他的概念一样,很是模糊。理智和情感同时工作,相互沟通,然后做出决定,最大程度上确保结果是积极的。尽管有的时候做出的决定会让你后悔,可是那只不过是人类学习的必要过程之一。
亲爱的欧律狄刻,请你不要哭泣
既然情感对于我们的行为如此至关重要,不出所料,人类会有意识或无意识地通过调控其他人的情感以改变其行为。人际情感调节(interpersonal emotion regulation)在社交生活中起到着很重要的作用:共情者为了共情对象的身心健康或为了自己的利益去调节共情对象的情感状态(Zaki, J. 2020)。他们在调节的过程中会采用不同的方式,例如以共情对象的角度引导情感改变的合作式(cooperative);或者是强制改变共情对象情感的家长式(paternalistic)。
各种政治运动的组织者会发表令人激奋的言论,这让愤怒的听众更有可能参与有风险的活动,所以组织者更有可能可以改变他们的现状。相反的,如果恐惧情绪被激发,人们会更倾向于避免风险,变得更加保守,来保护自己免受伤害。显而易见,在生活中,人们都会通过改变他人情感来达到自己的目的。
但是人际情感调节需要一个前提,那就是拥有共情能力。在猴子的运动前皮层(premotor cortex)F5区域发现的镜像神经元引出了人们对共情能力的思考。这些镜像神经元会在猴子做出一个动作时或在猴子看到其他猴子或者实验者做出同样的动作时激活。许多研究发现这些神经元的功能不仅仅局限于观察,它们对动作的理解,激发和学习起到很重要的作用。
在学习方面,现在的理论是猴子观察其他猴子做一个动作,然后那些被激活的镜像神经元在它自己去做类似的动作时可以再次被激活,而动作的结果会被反馈到大脑,允许大脑去继续调整,继续学习,直到这个动作被学会为止(Rizzolatti et al. 2004)。对于动作的理解方面,镜像神经元会在猴子看到实验者拿食物的动作时被激活,但是如果食物被挡住了,猴子只能看到同样的动作时,这些神经元依旧会被激活。这意味着这些镜像神经元帮助猴子理解动作而非单纯观看动作(Umilta et al. 2001)。
相似的镜像神经元在人类大脑中也同样存在,而他们的位置不仅仅局限于F5区域。不光如此,这些镜像神经元还与情感相关的脑岛连接。如果将镜像神经元的功能延伸到共情能力,观察共情对象的表情或情感表达可以激活人类镜像神经元,因此可以激发自己相关的情感经历,产生共情。但是值得注意的是,虽然镜像神经元在共情能力中是一个比较重要的组成部分,这并不意味着它们可以解释共情的所有过程,因为共情很多时候是不需要共情者看到共情对象的表情的。
回到最开始
自我意识的困难问题(hard problem of consciousness)提出了如果简单的输入输出就可以完成人们日常面对的任务,那么体验这些外界信息似乎变得多余。对于情感而言,经历一份情感和感受知觉是密不可分的,因为我们的大脑会把所有信息,甚至在你没有意识的情况下,分类成“令人愉悦的/好的”和“令人厌恶的/坏的”。若想处理外界的信息,它们的性质就必须用一种统一的标准来对比,而这个标准就是情感。情感体验是知觉的必然结果。
至于其他经历,例如视觉体验,我认为也是信息总和的一种方式。大脑需要把所有在一个空间里的事物信息都反馈到同一个单元,所以他们能够被同时处理,而在这种总和过程中我们有了知觉和体验。换句话来说,体验不是最终可有可无的过程,而是大脑要同时处理多种信息的方法,没有了体验,我们便没有了总和信息的能力。
相比之下,原核生物里的感光体就没有办法形成体验,因为它们不会总和各种有关周围环境的信息(例如物体的形状,材质),它们只会反馈光的强度,并且在被激活的时候直接导致细胞内的输出,对不同的光强度做出不同的反应。
当信息很简单而且信息量很小的时候,体验是不必要的,因为每一个输入信息不需要被拼凑在一起做出决定,生命体此时只需简单地做到输入输出就够了。对所有的生物来说,接受信息的物理过程可以不导致体验或自我意识,但是复杂的信息处理就需要体验来综合所有信息以做出决定。
参考文献
Kassam, K. S., Markey, A. R., Cherkassky, V. L., Loewenstein, G., & Just, M. A. (2013). Identifying Emotions on the Basis of Neural Activation. PLoS ONE, 8(6), e66032. Retrieved November 2, 2020, from 10.1371/journal.pone.0066032
Löken, L. S., Wessberg, J., Morrison, I., McGlone, F., & Olausson, H. (2009). Coding of pleasant touch by unmyelinated afferents in humans. Nat Neurosci, 12(5), 547–548. Retrieved November 2, 2020, from 10.1038/nn.2312
Miskovic, V., & Keil, A. (2014). Escape from harm: linking affective vision and motor responses during active avoidance. Soc Cogn Affect Neurosci, 9(12), 1993–2000. Retrieved November 2, 2020, from 10.1093/scan/nsu013
Olausson, H., Lamarre, Y., Backlund, H., Morin, C., Wallin, B. G., Starck, G., Ekholm, S., et al. (2002). Unmyelinated tactile afferents signal touch and project to insular cortex. Nat Neurosci, 5(9), 900–904. Retrieved November 2, 2020, from 10.1038/nn896
Rizzolatti, G., & Craighero, L. (2004). THE MIRROR-NEURON SYSTEM. Annu. Rev. Neurosci., 27(1), 169–192. Retrieved October 19, 2020, from 10.1146/annurev.neuro.27.070203.144230
Small, D. M., Gregory, M. D., Mak, Y. E., Gitelman, D., Mesulam, M. M., & Parrish, T. (2003). Dissociation of Neural Representation of Intensity and Affective Valuation in Human Gustation. Neuron, 39(4), 701–711. Retrieved November 2, 2020, from 10.1016/s0896-6273(03)00467-7
Todd, R. M., MacDonald, M. J., Sedge, P., Robertson, A., Jetly, R., Taylor, M. J., & Pang, E. W. (2015). Soldiers With Posttraumatic Stress Disorder See a World Full of Threat: Magnetoencephalography Reveals Enhanced Tuning to Combat-Related Cues. Biological Psychiatry, 78(12), 821–829. Retrieved November 2, 2020, from 10.1016/j.biopsych.2015.05.011
Todd, R. M., Miskovic, V., Chikazoe, J., & Anderson, A. K. (2020). Emotional Objectivity: Neural Representations of Emotions and Their Interaction with Cognition. Annu. Rev. Psychol., 71(1), 25–48. Retrieved October 19, 2020, from 10.1146/annurev-psych-010419-051044
Umiltà, M. A., Kohler, E., Gallese, V., Fogassi, L., Fadiga, L., Keysers, C., & Rizzolatti, G. (2001). I Know What You Are Doing. Neuron, 31(1), 155–165. Retrieved November 2, 2020, from 10.1016/s0896-6273(01)00337-3
Zaki, J. (2020). Integrating Empathy and Interpersonal Emotion Regulation. Annu. Rev. Psychol., 71(1), 517–540. Retrieved October 19, 2020, from 10.1146/annurev-psych-010419-050830
本文来自微信公众号:神经现实(ID:neureality),作者:杞人,编辑:阿莫東森