
本文来自微信公众号:集智俱乐部(ID:swarma_org),作者:郭瑞东,审校:张澳,编辑:邓一雪,题图来自:视觉中国

论文题目:Engineering self-organized criticality in living cells
论文地址:https://www.nature.com/articles/s41467-021-24695-
一、自组织临界其实并不陌生
夏日孩子们在沙滩上玩耍,常常堆出越来越大的沙丘。当沙丘堆积时,沙丘斜坡角度会达到一个临界点,一旦到达这个临界点,继续堆积沙子会导致沙丘以雪崩的形式向下滑落。在滑落前的最高点,沙堆处于有序与失稳之间的临界状态。
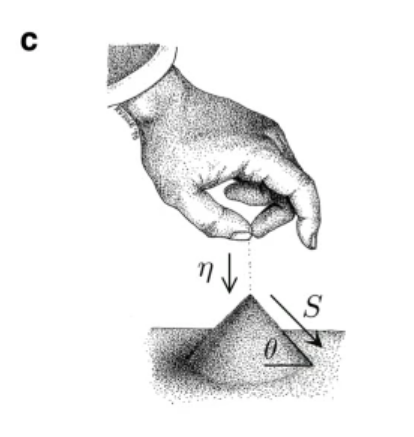
在沙堆模型中,通过向沙堆以速率η添加沙砾,沙堆倾角θ变大,刚开始只有少量沙粒落下,但随着倾角θ接近临界角θc,沙粒数量S迅速增加,直至θ=θc时,再加入的沙子会造成雪崩式的崩塌,造成沙墩倾角θ变小,重新回到临界态,下图描述了其反馈回路。
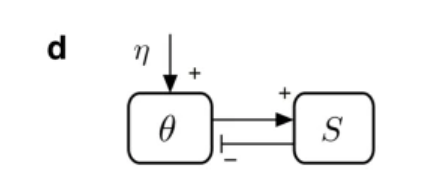
自组织临界性,由 Per Bak 、汤超、Wiesenfeld在1987年提出,该原理指出,系统元素之间的微小相互作用,会在没有外界介入的情况下,自然地到达临界点。在这个临界点上,一件微小的事件可以产生灾难性的反应。自组织临界性能在金融市场、交通体系等复杂系统中观察到。
细胞中的生化反应,大脑中的神经元激活,计算机的通讯网络,密封群落,这些系统都处在混乱和失稳之间的临界态。生活在临界点附近具有适应性优势,例如能更高效地处理信息,以迅速响应环境变化。因此,一个系统可以通过所谓的自组织临界性(self-organized criticality ,SOC)使自己稳定在临界点附近。
二、单细胞层面的自组织临界态
临界态作为多细胞认知系统的关键,能否出现在单细胞中?现有研究指出,在细胞水平上,酶网络可能处于临界态,以提升其对刺激的适应能力。该研究构建了一个简单的双基因网络,通过设计细胞内部基因表达和调控参数之间的相互作用,成功地让活细胞(细菌)进入了自组织临界态,找出了在构成自组织临界态所需的最小条件。同样的方法也可用于实现相关功能,如改善抗结直肠肿瘤药物的输送。
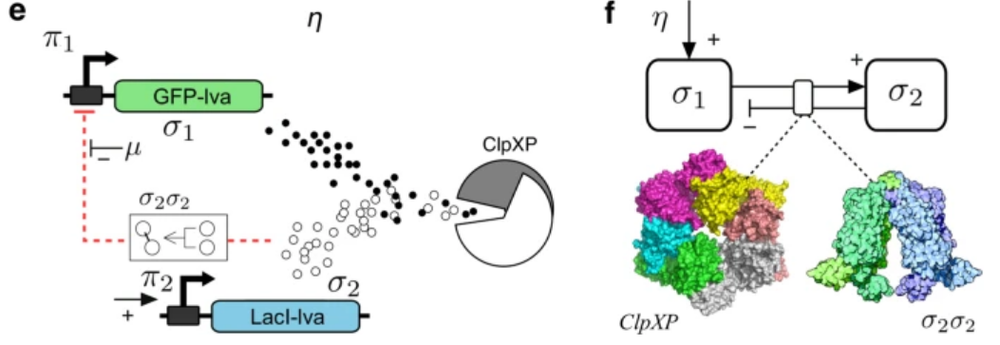
该研究中的大肠杆菌包含GFP-lva基因,该基因会被表达为蛋白σ,而蛋白σ又会被细胞内的蛋白水解机制(ClpXP)降解,从而影响GFP-lva基因表达量。其调控机制如下图所示:
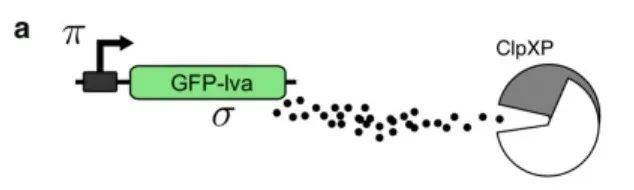
基因表达率η(横轴变量)较高时,细菌会产生过多的蛋白质,处于拥挤状态(下图中浅绿色的部分),而如果基因表达率较低,则细菌处于自由状态。通过调整基因表达率η,可确定划分临界状态与拥挤状态的临界表达率ηc,此时细菌处于自组织临界态,在该位置,任何一点小的变化都会对环境中蛋白质σ的浓度有显著影响。
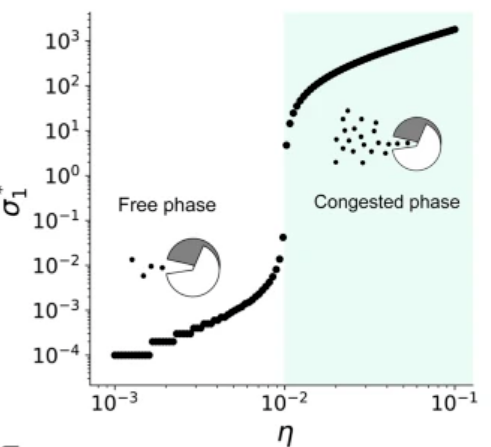
在大肠杆菌中,引入另一个同样竞争ClpXP的蛋白质,会促成一个类似沙堆模型的反馈回路,从而形成自组织临界态,下图是该研究中双基因组成的调控模型示意图。
三、自组织临界对合成生物学的意义
自然界中普遍出现的自组织临界现象,隐藏着生物之所以能高效且稳定运行的秘密。酶网络在底物输入速率与酶网络的处理能力相平衡时,呈现临界态,从而形成适应性优势。但对于达成临界态所需的最小条件仍不明确,该研究指出了实现临界态所需的基因网络中的最小模体。
生物计算指通过活细胞中的生化反应来进行信息处理任务,该研究设计的处在临界态的细胞,能够放大生物计算中的信号幅度,迅速形成雪崩式的信号传导,从而增强生物计算相关应用发展。由于多细胞认知及群体智慧都涉及临界态,该研究可支持未来相关领域的研究。同时,该研究对三维空间中细胞内部生化活动的物理建模也有所帮助。
本文来自微信公众号:集智俱乐部(ID:swarma_org),作者:郭瑞东