
大脑是神经系统最高级的部分。2013年,美国NIH启动“美国脑计划”。随着技术的不断更迭,2017年,NIH进一步启动BICCN(BRAIN Initiative Cell Census Network)项目,这是脑计划的重要组成部分,旨在对人类、非人灵长类动物和小鼠大脑中的不同细胞类型进行全面识别和归类。
2021年10月6日,BICCN在Nature重磅发布16篇文章介绍大脑运动皮层细胞图谱;2023年10月13日,又在Science系列期刊连发21篇论文,揭示了迄今最全面的人类脑细胞图谱。
就在上周,据Science发文仅两个月后,BICCN又有新动作,在Nature期刊同期发表了10篇研究论文,这10篇论文利用多种技术包括scRNA-seq、snRNA-seq、snm3C-seq、MERFISH(Multiplexed error-robust fluorescence in situ hybridization)、Slide-seq、STARmap、SMART-seq、snmC-seq、snATAC-seq分析了小鼠大脑中总计约3200万个细胞,鉴别出约5300个细胞类型,从而提供了迄今为止最全的小鼠完整大脑细胞类型的特性描述和分类。
这些论文的研究团队来自艾伦脑科学研究所(Allen Institute)、哈佛大学、索尔克生物研究所(Salk Institute for Biological Studies)、博德研究所(Broad Institute)、加州大学圣地亚哥分校和加州大学伯克利分校等知名脑科学研究机构,论文通讯作者包括曾红葵、庄小威、何志刚、任兵、王潇、刘嘉、陈飞等多位华人学者。

▷图1:Nature特刊封面与完整小鼠大脑细胞类型图谱。图源:nature
过去对大脑的探测工作受限于选定区域,而此次的10篇论文突破了局限,提供了对整个小鼠大脑的详细调查。研究内容可分为三个主题:
(1)构建小鼠全脑转录组学与空间图谱;
(2)构建小鼠全脑脊髓投射神经元与脊椎动物视网膜神经元分类图谱;
(3)剖析小鼠全脑表观遗传学特征与基因调控元件。
这些发现对完整的哺乳动物大脑结构和组织,以及单个脑细胞和神经回路的功能提供了详尽的信息。此外,它们也为进一步研究哺乳动物大脑的发育和演化提供了工具,包括不同类型的细胞组织引起神经系统疾病的微观机制。接下来,追问将带领读者一起了解此次的重要研究成果。
一、构建小鼠全脑转录组学与空间图谱
▷图2:利用MERFISH对约1000万个细胞中的1100个基因成像,生成分子定义和具有空间分辨率的全小鼠大脑细胞图谱。图源:nature
(一)小鼠全脑高分辨率的转录组及空间细胞图谱
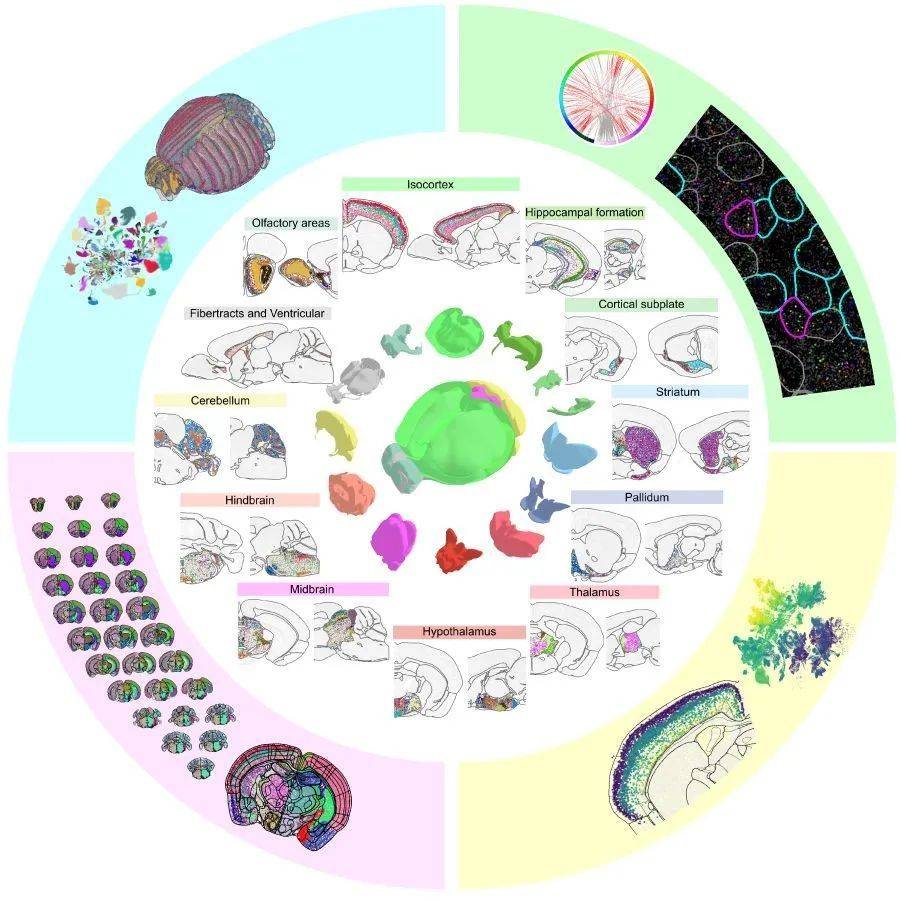
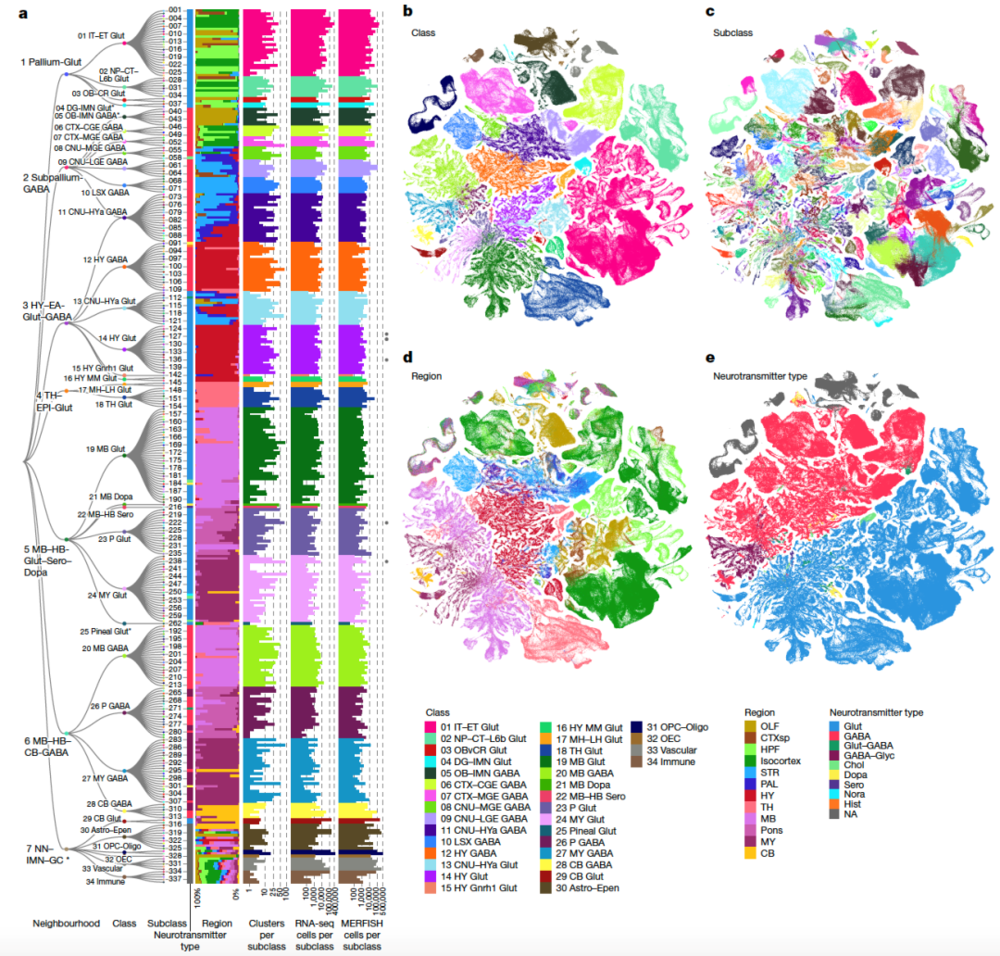
在该专题的旗舰论文中,美国艾伦研究所的曾红葵等研究人员描述了他们是如何从结合了约400万细胞的单细胞RNA测序和约430万细胞的空间转录组学数据中,创建出首个完整小鼠大脑高分辨率转录组细胞类型图谱的。
研究团队结合了两个海量数据集——约700万个细胞的单细胞RNA测序(scRNA-seq)数据和约430万个细胞的空间转录组学(MERFISH)数据,将小鼠全脑细胞分为34个大类(class),再进一步细分为338个子类(subclass),1201个超型(supertype),5322个细胞集群(cluster)。其中,神经元的细胞类型显示出惊人的多样性,共有5205个集群。
▷图3:小鼠全脑转录组细胞类型分类。图源:参考文献[1]
研究者指出,空间分辨率高是这份图谱的独特之处,由此该图谱可以将每种细胞的转录组特征与其空间位置高度对应,从而清楚地观察到大脑解剖结构与细胞类型之间的关系。
从图谱中可以观察到不同脑区细胞类型组织的独特特征,例如,在大脑的背侧和腹侧,细胞类型的组织存在明显差异:背侧区域含有的细胞类型较少,但高度分化;腹侧则包含了更多类型的神经元,但类型之间的关系较近。这种差异与不同结构的功能和演化有关。背侧部分包括皮质、丘脑和小脑等,主要执行生物体的适应性功能,如认知、感觉运动等,演化时间较晚,细胞类型在多样性上的拓展速度较快;腹侧包括下丘脑、中脑和后脑等较古老的结构,承担了进食、繁殖、新陈代谢平衡等更基本的生存功能,专用细胞类型和神经回路的变化就相对较小。
此外,这项研究还发现,不同细胞类型中神经递质和神经肽表达和共表达模式具有极为丰富的多样性和异质性。进一步研究表明,转录因子是细胞类型分类的主要决定因素。研究也确定了整个大脑各个部位决定每一种细胞类型的主要转录因子组合。
阅读原文:
Yao, Z., van Velthoven, C.T.J., Kunst, M. et al. A high-resolution transcriptomic and spatial atlas of cell types in the whole mouse brain. Nature 624, 317–332 (2023). https://doi.org/10.1038/s41586-023-06812-z
(二)小鼠全脑的分子与空间细胞图谱
哈佛大学庄小威实验室开发的多重容错性荧光原位杂交(multiplexed error-robust fluorescence in situ hybridization,简称MERFISH)技术在图谱创建中发挥了重要作用。这项技术可以让研究者在细胞或组织切片中同时检测多种RNA,从而绘制选定区域的基因表达空间图谱。
在这个由庄小威团队领衔的第二项研究中,研究者利用MERFISH对小鼠全脑约1000万个细胞中超过1100个基因进行了研究,绘制了一张超过5000个具有不同转录背景的细胞簇(归属于超过300个主要细胞类型)的高清分子与空间图谱(图4)。
过程中,研究团队将MERFISH与单细胞RNA测序数据结合进行成像,放入艾伦研究所的小鼠全脑通用坐标框架中,创建出单细胞空间图谱(每个细胞都有全转录组表达谱),可以系统地定量分析各个脑区的细胞类型和组成,推断细胞子类之间的特异性相互作用,预测配体-受体之间的分子关系,以及细胞-细胞相互作用的功能。
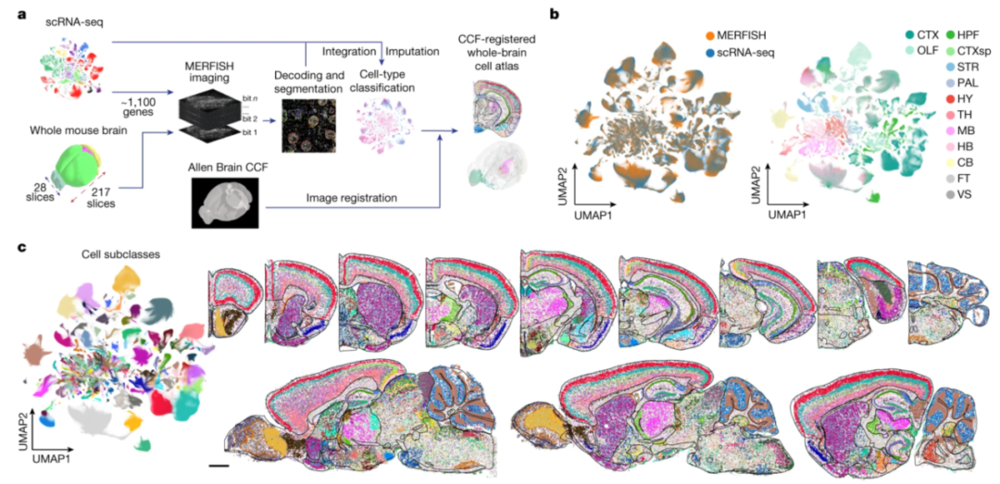
▷图4:构建全鼠脑细胞图谱的工作流程。图源:参考文献[2]
阅读原文:
Zhang, M., Pan, X., Jung, W. et al. Molecularly defined and spatially resolved cell atlas of the whole mouse brain. Nature624, 343–354 (2023). https://doi.org/10.1038/s41586-023-06808-9
(三)成年小鼠大脑的分子细胞架构
论文集的第三篇来自麻省理工学院和哈佛大学博德研究所。研究采用了不同但互补的方法,将新开发的空间转录组学技术Slide-seq与高通量单核RNA测序相结合,以440万个10微米像素的分辨率量化全基因组表达,使细胞类型能够系统地定位到单个大脑区域(图5)。这揭示了小鼠全脑各个解剖结构中的细胞类型组成,包括过去关注较少的中脑、脑桥、延髓和下丘脑。研究人员估计,分析已涵盖小鼠大脑中约90%的细胞类型。
研究发现,在进化上更为古老的大脑区域,特别是中脑、后脑和下丘脑,它们存在的细胞类型多样性异常惊人。分析结果表明,在中脑和下丘脑内发现的细胞类型比整个端脑内发现的还多。此外,在全脑分布上,具有长程投射的神经元的分布比局部中间神经元的分布更具区域特异性;非神经元类型的细胞如星形胶质细胞、外胚层细胞、伸展细胞、血管软脑膜细胞等,也具有非常明显的空间特异性,这表明了神经元与胶质细胞、胶质细胞与血管之间的特异性相互作用。
为了促进对相对未被充分研究的细胞类型功能的更多研究,研究团队通过在线开放资源的形式提供了这些数据。
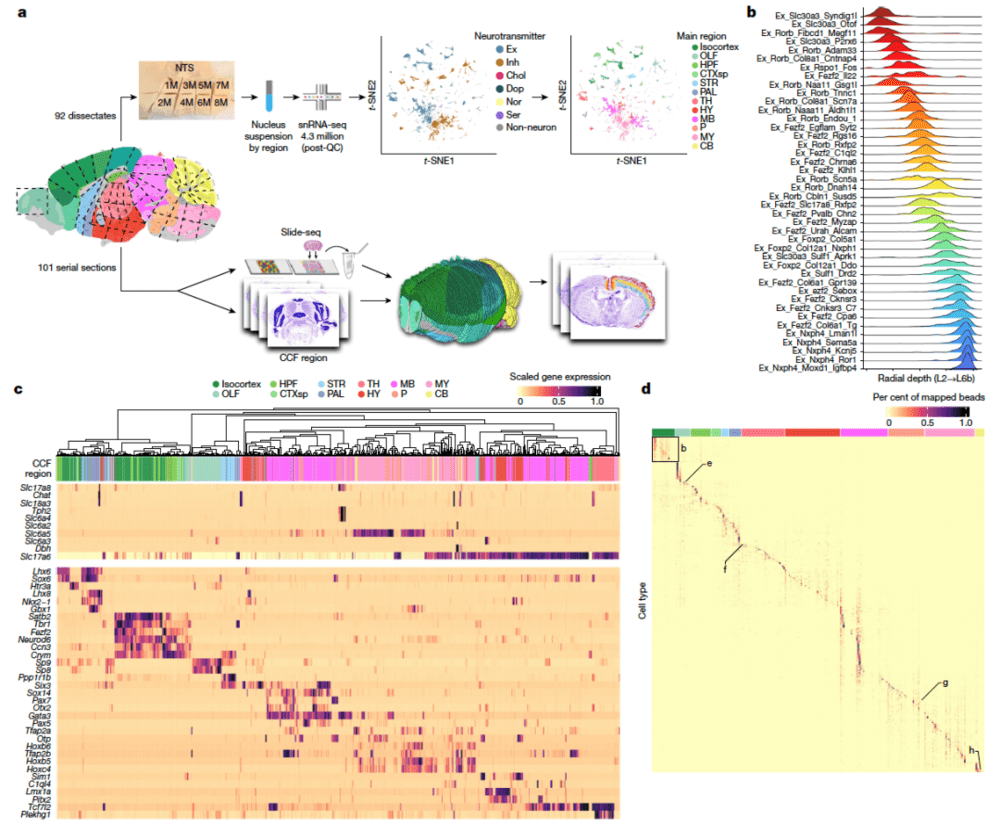
▷图5:利用全脑snRNA-seq和Slide-seq数据集绘制细胞类型的空间分布图谱。图源:参考文献[3]
阅读原文:
Langlieb, J., Sachdev, N.S., Balderrama, K.S. et al. The molecular cytoarchitecture of the adult mouse brain. Nature 624, 333–342 (2023). https://doi.org/10.1038/s41586-023-06818-7
(四)小鼠全脑高分辨率单细胞空间转录组图谱
第四篇论文由博德研究所王潇团队联合哈佛大学刘嘉团队共同发表(注:该论文已于2023年9月27日发表)。研究者采用原位空间转录组学技术STARmap PLUS,以194X194X345nm3体素尺寸在亚细胞分辨率水平上对成年小鼠全脑和脊髓中17个冠状切面和3个矢状切面组织切片中的1022个基因进行了检测,并使用细胞分割算法ClusterMap得到了109万个高质量空间分辨单细胞基因表达(图6)。
通过大规模的单细胞分析注释,该团队基于单细胞基因表达定义和注释了230个“分子细胞类型”并基于空间基因表达定义和注释了106种“分子组织区域”。在这一过程中,研究人员发现了部分端脑抑制性中间神经元亚型的脑区分布特异性。例如,纹状体中特有的中间神经元、嗅球的外网状层中表达多巴胺的中间神经元。
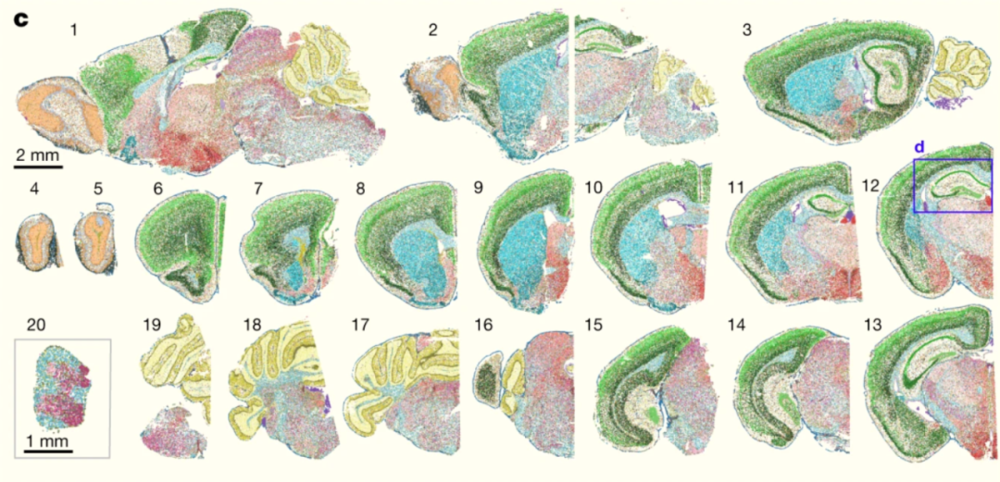
▷图6:小鼠中枢神经系统单细胞分辨率细胞类型空间图谱,230种细胞类型由彩色点标注。图源:参考文献[4]
基于空间上的分子表达,研究人员还补充和完善了小鼠大脑解剖学结构。例如,研究人员从分子表达和细胞分布的角度出发,划分小鼠大脑皮层的分区,并与传统解剖学的定义比较。一个有趣的发现是,解剖学上的压后皮层在小鼠大脑的前端和后端具有截然不同的“分子组织区域”组成;后端压后皮层在“分子组织区域”组成上与相邻的视觉皮层有着更高的相似性,这为理解压后皮层在视觉相关的行为和记忆中的功能提供了新的思路。
阅读原文:
Shi, H., He, Y., Zhou, Y. et al. Spatial atlas of the mouse central nervous system at molecular resolution. Nature 622, 552–561 (2023). https://doi.org/10.1038/s41586-023-06569-5
二、剖析小鼠全脑表观遗传学特征与基因调控元件
在论文集的第二个专题中,多个研究团队应用小鼠全脑转录组细胞类型图谱,分析了不同细胞类型的基因调控程序,并用于识别造成神经系统疾病和特性的遗传变异(图7)。
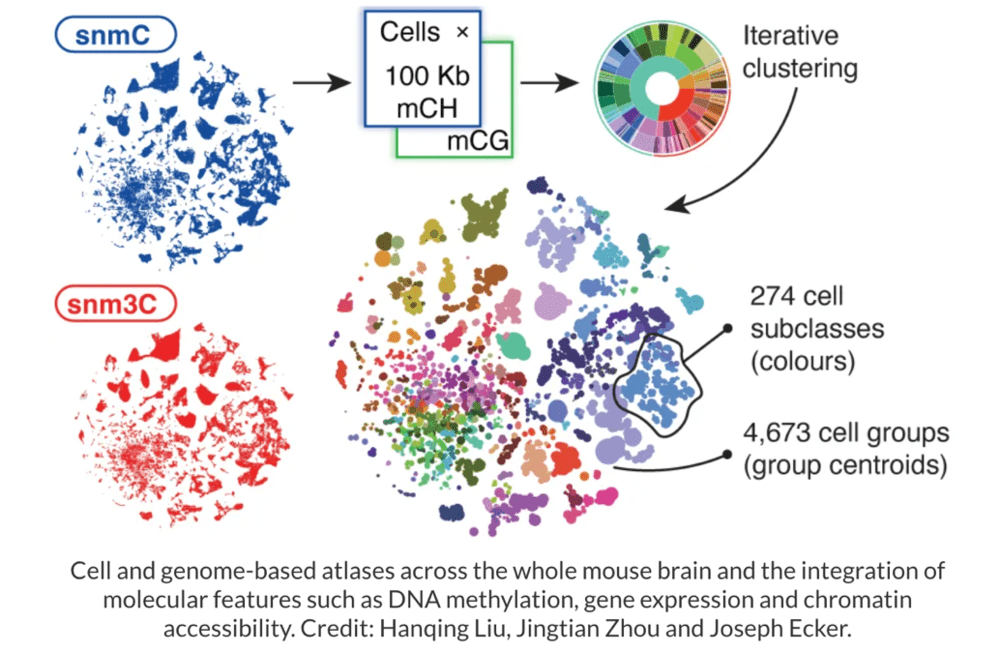
▷图7:整个小鼠大脑的细胞和基因组图谱,以及DNA甲基化、基因表达和染色质可及性等分子特征的整合。图源:nature
(一)小鼠大脑神经元表观组及远端投射图谱
大脑中的许多细胞类型通过远距离投射来调节细胞功能,还有许多特定细胞类型之间具有相互作用。加州大学圣地亚哥分校的Edward M. Callaway团队和索尔克生物研究所Joseph R. Ecker合作,通过Epi-Retro-Seq将单细胞表观基因组和细胞类型与小鼠全脑中32个不同区域的33034个神经元的长距离投射联系起来,这些神经元投射到24个不同的靶点(图8)。
这个数据集涉及大脑32个不同区域超过3万个大脑神经元。借助该数据集,研究者可以量化投射到不同目标脑区神经元之间的遗传差异,对分子细胞类型和它们的投射靶标进行注释,并且在投射富集的细胞类型中构建基因调控网络。
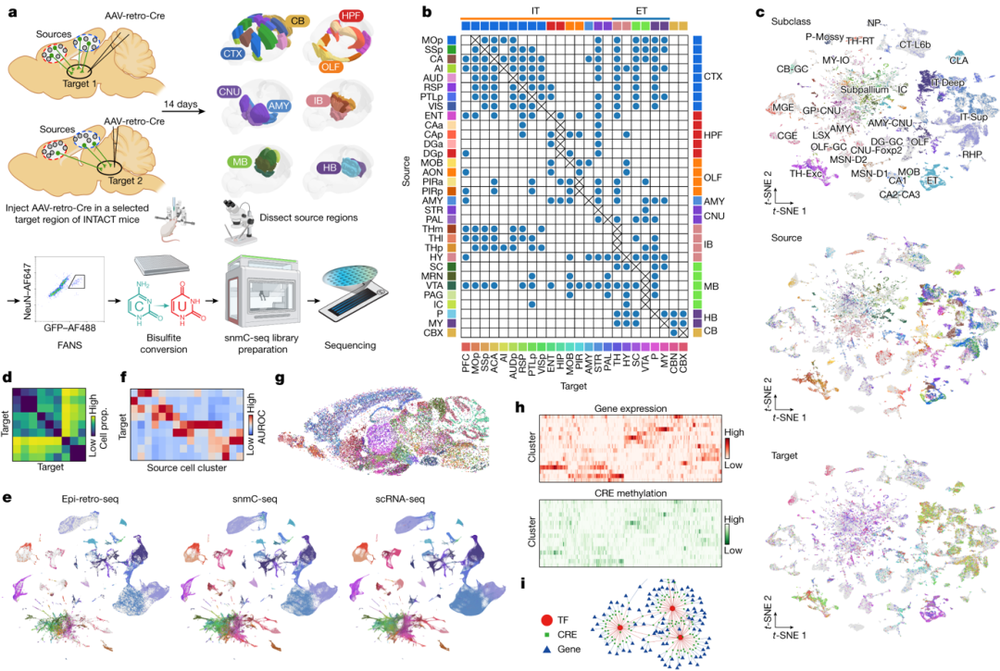
▷图8:全脑投射神经元的表观基因组图谱。图a.b表示用于逆行标记和表观遗传分析单个投射神经元的Epi-Retro-Seq工作流程。图源:参考文献[5]
阅读原文:
Zhou, J., Zhang, Z., Wu, M. et al. Brain-wide correspondence of neuronal epigenomics and distant projections. Nature 624, 355–365 (2023). https://doi.org/10.1038/s41586-023-06823-w
(二)成年小鼠全脑单细胞DNA甲基化和三维多组学图谱
胞嘧啶DNA甲基化对大脑发育非常关键,并与各种神经系统疾病有关。索尔克生物研究所Joseph R.Ecker团队尝试在空间背景下理解大脑的DNA甲基多样化。借助单细胞DNA甲基化测序(snmC-seq3)和甲基化联合三维基因组测序(snm3C-seq)技术,首次构建了完整的大脑细胞类型及其基因调控的分子图谱(图9)。
研究人员一共从成年小鼠大脑的117个解剖区域解析了超30万个甲基组细胞和17万个甲基组联合三维基因组细胞。通过迭代聚类和整合分析,联合了脑计划中的转录组数据集(10XRNA)以及基因组可及性数据集(snATAC-seq),构建了一个基于甲基化的细胞分类图谱,包括4673个细胞群组和274个多组学注释的亚类。
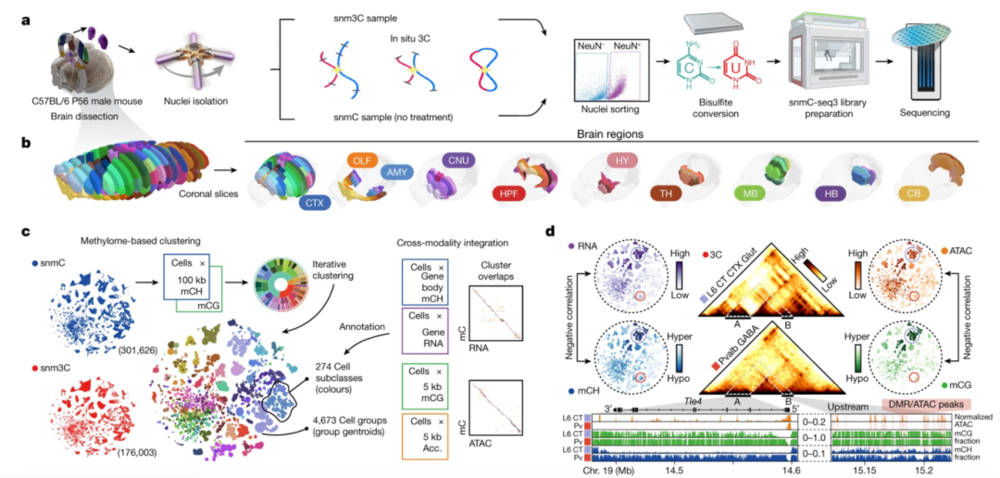
▷图9:snmC-seq3 + snm3C-seq技术工作流程示意图。图源:参考文献[6]
研究人员系统比较了全脑单细胞甲基化组学数据集与空间转录组学(MERFISH)数据集的关联性,结果表明,不同细胞的基因组甲基化模式与其在大脑中的空间位置高度相关。通过比较数百种不同的神经元亚类,研究人员还发现三维基因组结构也与DNA甲基化高度相关,尤其是在众多与神经细胞功能相关的长基因周围。这些全脑单细胞DNA甲基化组图谱可以为了解小鼠大脑细胞空间、基因组调控多样性提供大量资源。
阅读原文:
Yao, Z., van Velthoven, C.T.J., Kunst, M. et al. A high-resolution transcriptomic and spatial atlas of cell types in the whole mouse brain. Nature624, 317–332 (2023). https://doi.org/10.1038/s41586-023-06812-z
(三)成年小鼠大脑单细胞染色体可及性图谱
由加州大学圣地亚哥分校任兵团队绘制的成年小鼠大脑中顺式作用DNA元件(candidate cis-regulatory elements, cCREs)综合图谱,通过分析230万个单细胞ATAC-seq数据而生成。整个图谱包括约100万个cCREs,以及它们在1482个不同脑细胞群体中的染色质可及性。与早期该类研究相比较,新研究为小鼠基因组新增加了44.6万多个cCREs(图10)。
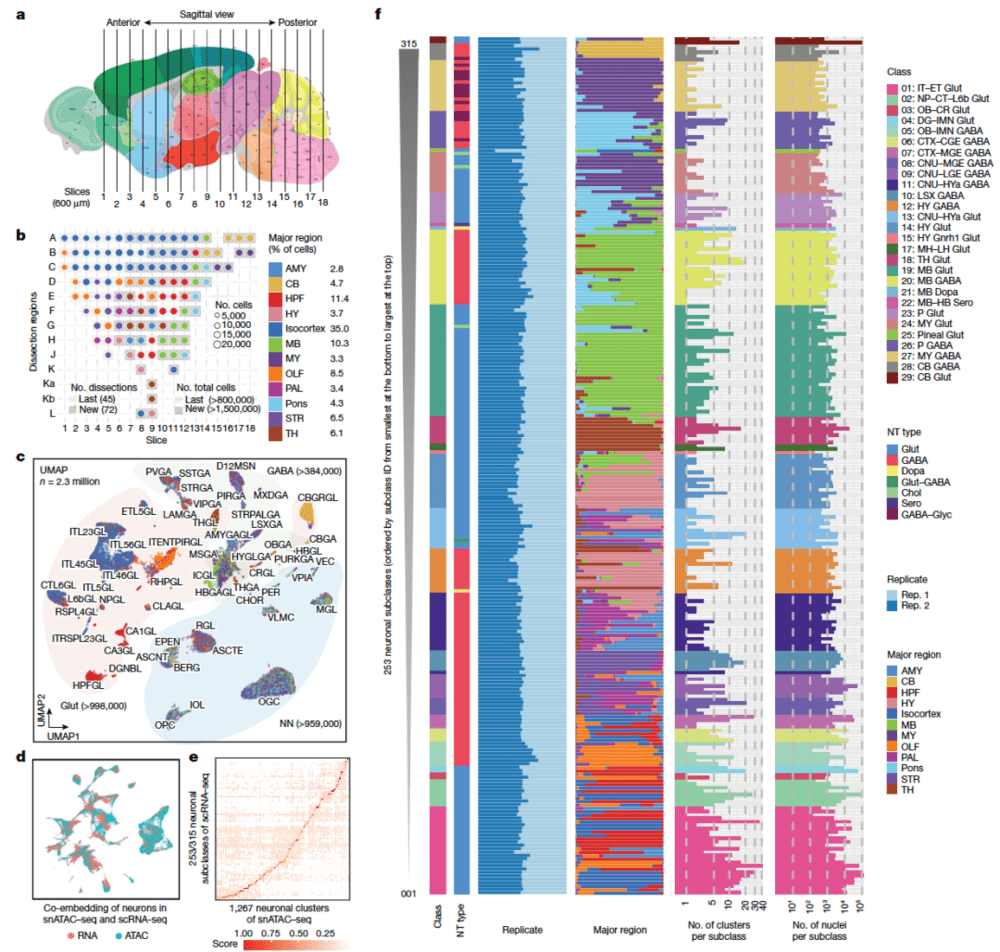
▷图10:成年小鼠全脑染色体可及性的单细胞分析。图源:参考文献[7]
根据该染色体可及性图谱,研究发现转座子(transposable elements, TEs)在新的调控程序和神经元多样性中具有潜在作用。例如,较其他小鼠脑细胞,在部分皮层兴奋性神经元中,研究人员观察到小鼠特异的cCREs更为显著地在TEs中富集。通过整合单细胞ATAC-seq数据和单细胞RNA-seq数据,研究人员刻画了260多个小鼠脑细胞亚型的基因调控网络,并利用深度学习工具,训练了一个可以仅根据DNA序列预测不同脑细胞类型中cCREs的染色质可及性的模型。这项工作为分析小鼠和人类全脑的细胞类型特异的基因调控程序提供了坚实的数据支持。
阅读原文:
Zu, S., Li, Y.E., Wang, K. et al. Single-cell analysis of chromatin accessibility in the adult mouse brain. Nature624, 378–389 (2023). https://doi.org/10.1038/s41586-023-06824-9
(四)多组学揭示哺乳动物初级运动皮层的基因调控
在该专题的最后一项研究中,任兵团队和Joseph R.Ecker团队强强联合,利用单细胞多组学方法研究了人类、猕猴、小鼠初级运动皮层的基因调控程序,从超过20万个细胞中获得了基因表达、染色体可及性、DNA甲基组和染色体构象图谱(图11)。这些数据显示,转录因子表达差异与物种特异性表观基因组相对应。
研究人员发现,保守和分化的基因调控特征可以在三维基因组的演化中得以反映。在皮层细胞中,TEs占据了人类约80%的cCREs。利用机器学习手段,研究人员在不同物种中开发了可以预测顺式调控元件的预测因子,该模型揭示了从啮齿类到灵长类基因组调控规则的高度保守性。表观遗传的保守性结合序列相似性有助于挖掘功能性cCREs,帮助我们理解神经系统疾病的遗传变异机制。例如,该研究还发现了与多发性硬化、厌食和烟瘾相关的遗传变异中的保守特征。
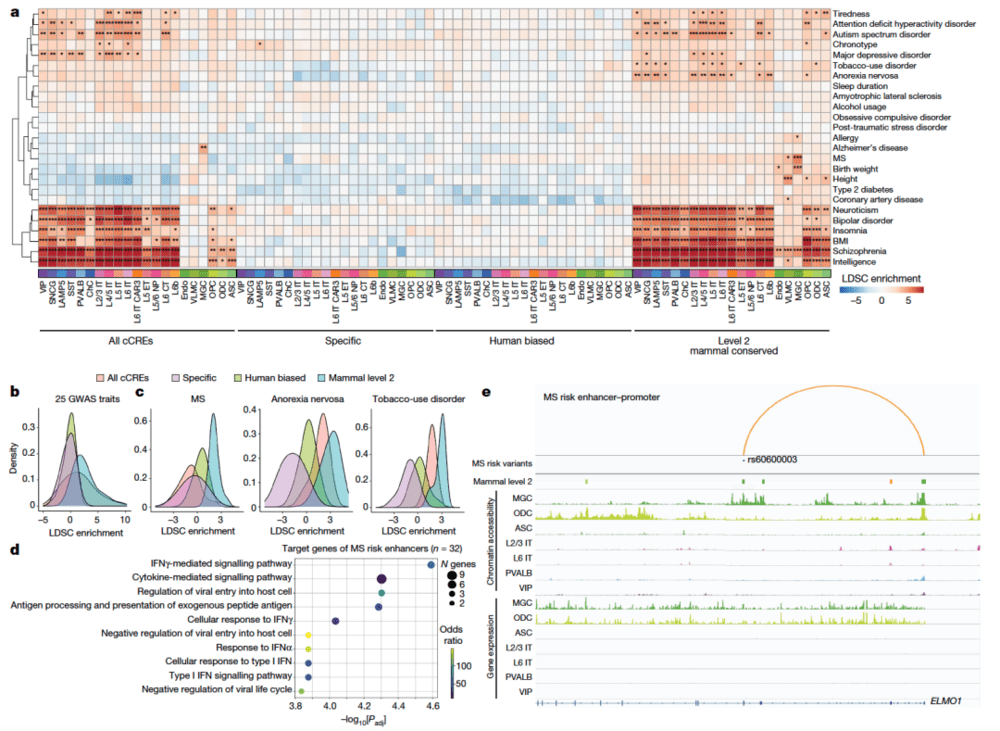
▷图11:利用表观遗传学保守特征解释与神经系统疾病风险相关的非编码变异。图源:参考文献[8]
阅读原文:
Zemke, N.R., Armand, E.J., Wang, W. et al. Conserved and divergent gene regulatory programs of the mammalian neocortex. Nature 624, 390–402 (2023). https://doi.org/10.1038/s41586-023-06819-6
三、小鼠全脑脊髓投射神经元与脊椎动物视网膜神经元分类图谱
基于对全脑细胞转录组的研究,论文集的第三个专题针对特定神经元类型,即小鼠脊髓投射神经元与脊椎动物视网膜神经元图谱进行了探索。
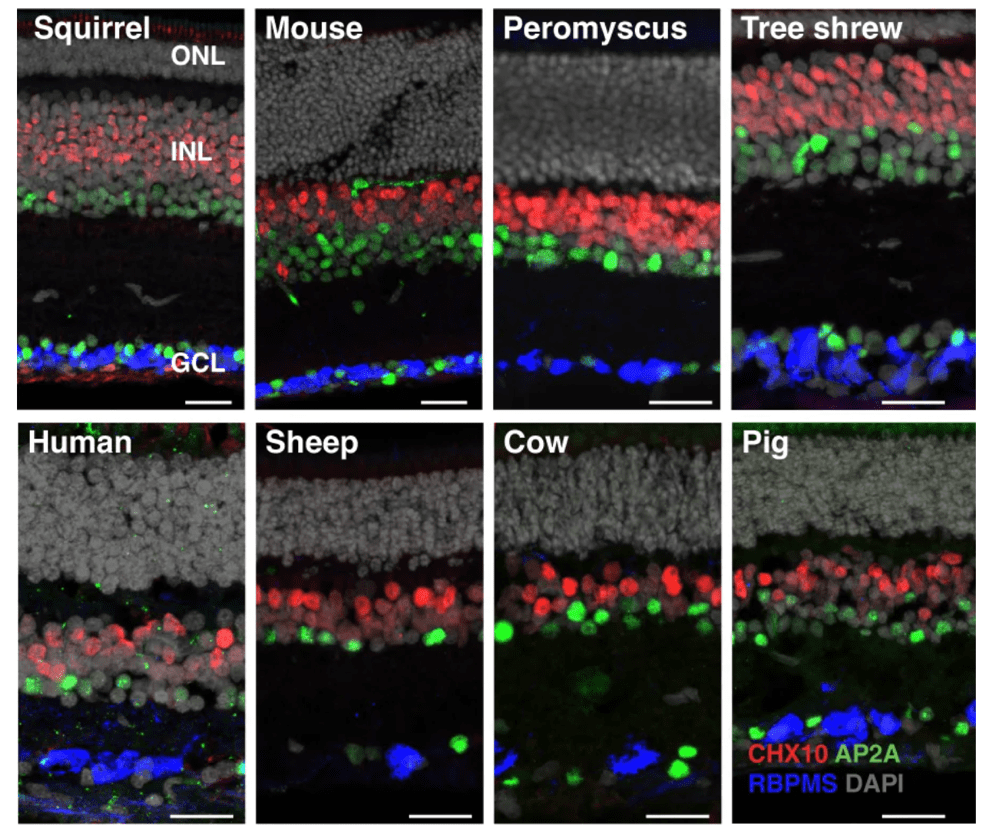
▷图12:对17种脊椎动物的比较转录组学分析揭示了灵长类动物中主导视觉处理的视网膜细胞类型的深度进化保守性,并确定了其他物种中的相应细胞。图源:nature
(一)小鼠全脑脊柱投射神经元的转录组分类
下行通路由直接从大脑各个区域投射到脊髓的脊柱投射神经元(spinal projecting neurons, SPNs)组成,SPNs将信号由大脑传递至脊髓,将大脑的指令(“思想”)转化为身体行为的指令(“行动”)。这些SPNs不仅对自主和非自主运动至关重要,而且在感觉调节和自主功能(如血压、心率和恐惧反应)中也起到关键作用。然而,对全脑SPNs的全面分子表征仍然不足。
哈佛医学院何志刚和艾伦研究所曾红葵合作,对65,002个SPNs进行了转录组分析,确定了3个群体、13个亚类和76种类型,遍及大脑皮层、下丘脑、中脑、桥脑背侧被盖体、桥脑网状结构和小脑深核,并将这些类型映射到整个小鼠大脑图谱中[1],以获得每种类型的精确空间位置(图13)。
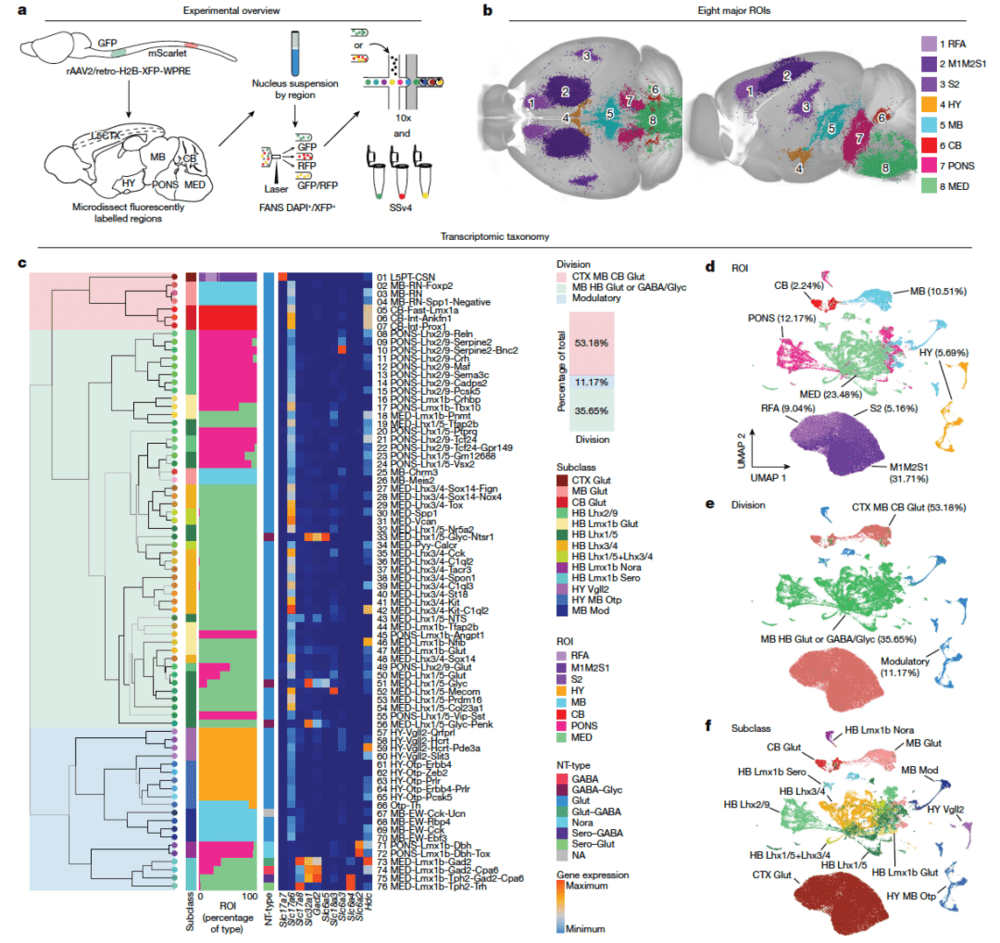
▷图13:全脑脊柱投射神经元的解剖学转录组图谱。图源:参考文献[9]
具体来说,该研究将全脑SPNs分为三大群体。
(1)来自皮质、红核和小脑的具有分子同质性的兴奋性SPNs,这群神经元可能适用于脑于脊髓的“点对点”信息传递;
(2)网状核分子异质性SPNs种群,将信息传递至脊髓多个区域,适合传递与整个脊髓活动相关的指令;
(3)在下丘脑、中脑和网状核中表达缓慢作用的神经递质和/或神经肽的调节神经元,用于脑脊髓信号的“增益”。
这项研究揭示了全脑SPNs的全面分类,并深入探讨了SPN在影响大脑对身体功能控制中的功能组织。
阅读原文:
Winter, C.C., Jacobi, A., Su, J. et al. A transcriptomic taxonomy of mouse brain-wide spinal projecting neurons. Nature 624, 403–414 (2023). https://doi.org/10.1038/s41586-023-06817-8
(二)脊椎动物视网膜中神经元细胞类型及演化
在该主题的另一篇论文中,加州大学伯克利分校Karthik Shekhar和哈佛大学Joshua R. Sanes研究团队合作,共同解开了利用细胞类型图谱窥视动物眼睛的演化之谜。
视网膜是眼睛直接与脑部相连的结构,也是视觉感知的起点。不同动物的视觉差异巨大,但新研究却指出,不同脊椎动物视网膜的细胞类型有着惊人的一致性。研究人员首先生成并比较了包括人类在内17种脊椎动物的视网膜单细胞转录组图谱,这些物种跨越了广泛的动物类型,但6种视网膜细胞类型(光感受器、水平细胞、双极细胞、无长突细胞、视网膜神经节细胞和穆勒胶质细胞)有着高度分子保守性,但随着物种进化距离也存在转录差异(图14)。基于保守的基因表达程序,众多细胞类型在不同物种间共享。
这些发现提供了一种视网膜进化的可能性:早在2亿年前,所有哺乳动物最后共同祖先的视网膜,就已经具有与现代哺乳动物相媲美的复杂性;其中一些细胞类型的历史,甚至可以追溯到4亿多年前所有脊椎动物的共同祖先。
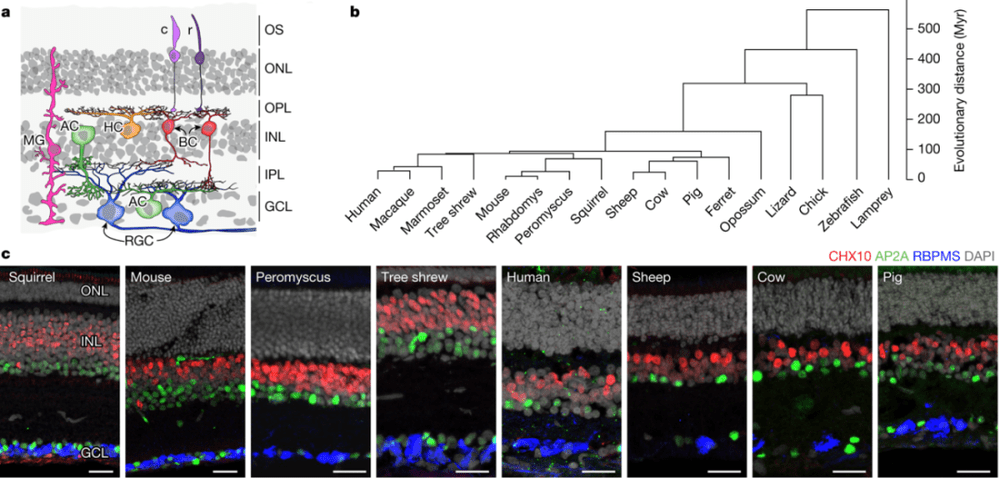
▷图14:脊椎动物视网膜的切片示意图,显示其六个主要细胞类别:光感受器(包括视杆细胞(r)和视锥细胞(c))、水平细胞(HC)、双极细胞(BC)、无长突细胞(AC)、视网膜神经节细胞(RGC)和穆勒胶质细胞(MG)。图源:参考文献[10]
这项研究还为一类视网膜神经节细胞(RGC)的起源提供了全新的认知。侏儒视网膜神经节细胞(midget RGC)占据了人类RGC细胞的80%以上,此前被认为源于灵长类动物,它们为人类高度灵敏的视力提供了基础。
但基于对小鼠脑图谱的对比研究发现,小鼠视网膜中也有与侏儒RGC同源的细胞alpha RGC。与人类的侏儒RGC相比,alpha RGC的细胞体更大,但数量更少,仅占小鼠RGC的2%~4%。追踪它们的进化表明,它们可能随着视觉皮层的扩大而变得更小和更多,而视觉皮层是人类视觉处理的主要中心。这种对应关系表明,视网膜和皮层同时进化,为灵长类动物提供了高灵敏度的视觉。
阅读原文:
Hahn, J., Monavarfeshani, A., Qiao, M. et al. Evolution of neuronal cell classes and types in the vertebrate retina. Nature 624, 415–424 (2023). https://doi.org/10.1038/s41586-023-06638-9
四、BICCN2.0 的意义何在?
在过去的十年里,单细胞RNA测序的技术建立推动了细胞类型图谱的产生。到目前为止,多模式、高分辨率的图谱主要局限于大脑的特定部位,比如运动皮层。BICCN2.0图谱将单细胞RNA测序与高分辨率单细胞转录组学技术结合,极致详尽地展示出全脑而不仅仅是某个脑区有多少种脑细胞类型,以及它们在全脑中的比例和空间排列。小鼠是神经科学研究中最常用的脊椎动物实验模型,这张图谱为更好地理解人类大脑铺平了道路。细胞图谱还为开发新一代精确治疗大脑精神和神经疾病的疗法奠定了基础。
细胞图谱描述了小鼠大脑每个区域的细胞类型及其在这些区域内的组织结构。研究表明,一个区域中神经元类型的数量并不与该区域的大小或神经元总数成比例。不同的规则支配着整个大脑的神经元多样性,这种现象可能是因为大脑的每个区域都在不同的约束下进化。转录因子决定了细胞类型的分类。在这些观察的基础上,研究人员进一步探究了脊柱投射神经元与视网膜神经元的转录特征。
此外,通过剖析小鼠全脑表观遗传学特征与基因调控元件,图谱揭示了在不同细胞类型和物种中,遗传密码(细胞DNA和染色体的化学修饰)如何被读取和解释的规则。研究表明,基因序列和细胞类型特异性表观基因组之间的联系在物种中是保守的。高度保守的调控元件往往位于它们所调控基因的起始位置附近,而物种特异性调控元件则位于更远的位置。小鼠或人类细胞特有的调控元件经常与转座元件重叠,这表明转座子可能被用于微调特定细胞类型和不同物种的基因激活。通过将表观基因组图谱与大脑不同部位细胞类型之间的投射联系起来,研究表明表观基因组可以塑造不同大脑区域细胞类型之间的相互作用,以及单个细胞的遗传活动。
2021年,BICCN的研究人员在大约1%的小鼠大脑中绘制了这些细胞类型。这些基因组技术的进步使研究小组在两年内就能对成年小鼠大脑中90%的细胞类型进行分类。目前,美国国立卫生研究院已经启动了另一个为期五年的项目,名为“大脑倡议细胞图谱网络(BICAN)”,其目的是扩大技术规模,绘制人类和非人类灵长类动物以及小鼠大脑在器官发育过程中的脑细胞类型。
BICAN代表了一个创新性的倡议,它与另外两个重要项目并行发展:大脑跨尺度连接计划(BRAIN Initiative Connectivity Across Scales)和精确获取脑细胞的装备计划(Armamentarium for Precision Brain Cell Access)。这些项目共同致力于解析控制行为神经回路的基本原理,并为治疗人类脑部疾病开辟新途径,以期彻底革新神经科学的研究领域。
参考文献:
[1]Yao,Z., van Velthoven, C.T.J., Kunst, M. et al. A high-resolution transcriptomic and spatialatlas of cell types in the whole mouse brain. Nature 624, 317–332 (2023). https://doi.org/10.1038/s41586-023-06812-z
[2]Zhang, M., Pan, X., Jung, W. et al. Molecularly defined and spatially resolvedcell atlas of the whole mouse brain. Nature 624, 343–354 (2023).https://doi.org/10.1038/s41586-023-06808-9
[3]Langlieb, J., Sachdev, N.S., Balderrama, K.S. et al. The molecular cytoarchitecture of the adultmouse brain. Nature 624, 333–342 (2023).https://doi.org/10.1038/s41586-023-06818-7
[4]Shi, H., He, Y., Zhou, Y. et al. Spatial atlas of the mouse central nervoussystem at molecular resolution. Nature 622, 552–561 (2023).https://doi.org/10.1038/s41586-023-06569-5
[5]Zhou, J., Zhang, Z., Wu, M. et al. Brain-wide correspondence of neuronalepigenomics and distant projections. Nature 624, 355–365 (2023).https://doi.org/10.1038/s41586-023-06823-w
[6]Liu, H., Zeng, Q., Zhou, J. et al. Single-cell DNA methylome and 3D multi-omicatlas of the adult mouse brain. Nature 624, 366–377 (2023).https://doi.org/10.1038/s41586-023-06805-y
[7]Zu, S., Li, Y.E., Wang, K. et al. Single-cell analysis of chromatinaccessibility in the adult mouse brain. Nature 624, 378–389 (2023).https://doi.org/10.1038/s41586-023-06824-9
[8]Zemke, N.R., Armand, E.J., Wang, W. et al. Conserved and divergent gene regulatoryprograms of the mammalian neocortex. Nature 624, 390–402 (2023).https://doi.org/10.1038/s41586-023-06819-6
[9]Winter,C.C., Jacobi, A., Su, J. et al. A transcriptomic taxonomy of mousebrain-wide spinal projecting neurons. Nature 624, 403–414 (2023).https://doi.org/10.1038/s41586-023-06817-8
[10]Hahn, J., Monavarfeshani, A., Qiao, M. et al. Evolution of neuronal cell classes and typesin the vertebrate retina. Nature 624, 415–424 (2023).https://doi.org/10.1038/s41586-023-06638-9
原始论文集:
https://www.nature.com/immersive/d42859-023-00069-2/index.html
本文来自微信公众号:nextquestion (ID:gh_2414d982daee),作者:梓薇,编辑:韵珂