
记忆形成的奥秘一直是人们探索大脑的重要方向。我们都知道睡眠对于巩固白天的记忆有重要的作用,然而,在众多日常经历中,大脑如何筛选出值得记住的瞬间?
科学家们认为,大脑可能会为清醒状态下的经历打上标签,以便于后续的记忆巩固,这一步在分子层面被称作“突触标记”。早在上世纪末便提出了一种哺乳动物脑中记忆的主要细胞模型:长时程增强(long-term potentiation ),这种模型可以实现晚期LTP的突触特异性,从而巩固某些“记忆”[1]。关于大脑如何在经历的实时处理过程中做出选择,神经生理学方面仍然缺乏充分的理论支持。
在经过长时间的探索后,目前最吸引科学家眼球的神经机制是海马体中的清醒尖波涟漪(awake sharp wave ripples, SPW-Rs)。科学家们认为,也许就是清醒尖波涟漪在帮助我们从经验中选择有用的部分,来为未来做准备。之前的研究表明尖波涟漪对于记忆巩固有重要作用[2, 3]。其中经验的显著特征,如新奇性和奖励的大小,促进了清醒状态下多个方面的重放,并增强了记忆[4]。
基于这些先前的研究,纽约大学的神经科学家们专门设计了一项新的实验,以研究哪些事件会被清醒尖波涟漪选择,并在睡眠中被尖波涟漪反复重放。为了保证不被外部人为因素(如何奖励、新奇性)的混淆,他们将实验设置的极为简单:让小鼠在八字迷宫中自由探索,随机给予奖励。通过在小鼠海马背侧CA1区植入双侧硅探针,科学家们获取了小鼠在探索过程以及睡眠过程中数百个神经元同时记录的脉冲放电序列。
为了确定SPW-Rs是否在标记特定事件,需要分两步来解决:首先,我们需要解码出整个探索过程脉冲活动序列的连续试次信息,也就是时间维度的解码;随后,如果我们能从海马神经元活动中解码出它属于哪一段时间,我们便可以将其用于清醒状态与睡眠状态下的尖波涟漪活动,看它们是否有在编码同一段经历。

Yang, Wannan, et al. "Selection of experience for memory by hippocampal sharp wave ripples." Science 383.6690 (2024): 1478-1483.
技术革新:从海马细胞群体中解码出时间信息
让解码时间信息成为可能,依赖于分析方法上的两个突破:序列非负矩阵分解(seqNMF) 与统一流形的估计投影技术(UMAP)。
seqNMF是一种专门用于分析时间序列数据的技术。其主要目的是从复杂的时间序列中提取出重复出现的序列结构,例如随时间变化而显现的模式或序列。这一方法通过将原始时间序列数据构建为一个非负矩阵,然后将其分解成两个或多个非负矩阵的乘积来实现,其中一个矩阵代表发现的模式或序列,另一个则代表相应的系数或权重。
通过这种方式,seqNMF能够识别数据中重复出现的模式或序列,并确定它们出现的时间点和权重[5]。在神经科学领域,seqNMF经常被用来解析神经元群体活动中的时间模式,有效地将杂乱无章的神经元放电序列数据整理成有序的模式,从而映射出与小鼠在八字迷宫中行为相匹配的神经放电模式。
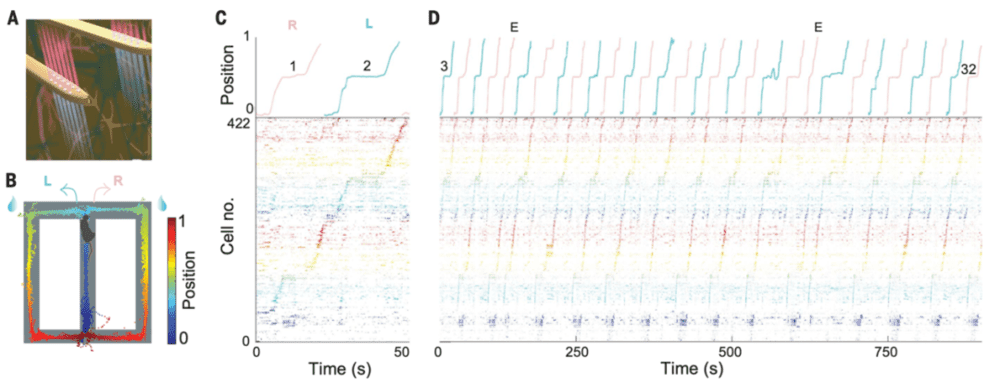
图1. A-D. 利用图A所示的记录手段,分析图B中小鼠运动所对应的神经活动。经过seqNMF排序后,神经元表现出了与小鼠实际运动相一致的发放模式。
而UMAP是一种用于降维和可视化高维数据的技术。它采用非线性的方法进行降维,工作原理类似于t-SNE,通过一种称为局部连通性的技术保留数据中的局部结构,同时尽量减少全局结构的失真。UMAP的一个显著优势在于,它能够在保留数据的内在结构的同时,最大程度上保持原始数据的全局布局,使其成为探索高维数据中隐藏模式和关系的强大工具[6]。
因此,UMAP为我们提供了一种新的方式来探索不同行为产生的放电序列之间的差异。通过将前文提到的高维神经元活动数据嵌入到低维流形空间,我们可以直观地观察到海马神经元群体活动所对应的隐藏空间与物理环境的拓扑结构之间的相似性,说明海马神经元群体能够编码小鼠运动的空间信息。更为有趣的是,如果我们按照实际运动路径在低维流形中将试次顺序用颜色进行编号着色,可以看到它们之间保持着对应的关系。这一观察结果在不同类型的迷宫和不同啮齿动物物种中都具有一致性。

图2.E-F. 将小鼠运动轨迹与神经元活动流形按照空间和时间分别编号着色,可以看到二者之间保持了拓扑上的对应关系。
在定性观察到时间序列信息在降维处理后仍能保持之后,我们需要进一步定量研究试次信息在状态空间中的表征方式。通过定义“试次块”概念,研究者将整个活动过程划分为不同的试次块序列,每个试次块内部的试次则被标记为第1至第5试次。随机选取1/10的试次作为测试集,使用kNN解码器来判断这些试次分别属于第1至第5试次中的哪一个。无论是在原始的高维空间还是在低维的UMAP流形中,该解码器均展示出了高准确率,这证明UMAP流形在低维空间中成功保留了原始高维空间中的试次序列信息。
在从神经元群体上成功解码出试次序列信息后,为了进一步探讨单个神经元对可解码性的贡献,研究人员绘制了每个神经元在不同试次中的调谐曲线,并观察到了各种试次间的变化模式,例如:在前后试次相同位置神经元的放电强度由强变弱或由弱变强,或是同一神经元在不同试次中编码不同位置。这些变化或差异是试次序列可解码性的关键来源。这种可解码性可能源自跨试次间的随机变化(即试次1至5之间的差异是随机的),也可能是试次间的结构性变化(即试次1至试次5之间按时间轴发生变化)。
为了验证这一假设,研究人员构建了一组模拟神经元,这些神经元具备与真实神经元相似的调谐曲线,但每个神经元的放电活动完全是独立的随机事件。使用与真实神经元相同的UMAP降维方法处理这组神经元后,由于调谐曲线的一致性,降维后的拓扑结构依然与真实环境相似。然而,在这些虚构神经元上,试次序列信息却消失了,这在可视化上表现为,当按照时间编号着色后,流形显得非常混乱。
另外研究人员还进行了一个与10-fold等价的交叉验证方法,不再是随机选取1/10的试次,而是每次选出一个完整的试次块,使用剩余的试次块来解码该试次块内的试次1-5的序列编号。在移除用于测试的数据后,研究人员使用剩下的所有试次块训练了一个kNN解码器,并得到了一个不包含某一试次块的低维流形。
随后,被取出的试次块被投影到这个低维流形上,以解码其试次序列。如果假设成立——数据的状态空间是沿着结构化的方式变化,那么作为测试集的试次块应该与其相邻试次块的距离最近,从而这些试次块中的试次1-5的判定准确性也应当最高。通过在真实数据和模拟神经元上进行此分析,结果显示,只有真实数据能够正确地将试次块解码到其前后相邻的试次块。
这一结果也在真实数据与打乱的数据之间的准确率比较中得到了证实。另外,虽然非位置细胞在降维后未能展示出与真实空间一致的拓扑结构,但在按时间编号着色后,其显示出的拓扑结构与时间变化保持了一致。研究中还发现,随着参与解码的神经元数量的减少,解码准确率迅速降低。
以上这些发现一方面表明,神经群体活动的试次间变化(漂移)并不能被单个神经元水平上的随机波动所解释。另一方面也证实了seqNMF和UMAP这两种分析方法在挖掘神经元群体时间信息编码上的有效性。接下来,需要将我们的目光转移到我们真正关心的海马尖波涟漪上去了。
重大发现:清醒与睡眠时期的尖波涟漪高度相关
我们已经知道,当动物主动在环境导航时,位置与序列编码与theta振荡密切相关;而当动物停下来饮水(获取奖励)时,theta波段活动便会停止,转而出现同步化的尖波涟漪活动。以往研究证实,在尖波涟漪中的脉冲序列能够重现物理空间中动物所经历的位置序列。现在利用seqNMF和UMAP对试次信息的编码能力,研究者们想要探索试次块序列是否也能通过尖波涟漪被重放。若此假设成立,我们就能同时揭示尖波涟漪信号中蕴含的空间和时间信息。
首先需要找出与重放有关的尖波涟漪:判断标准主要取决于该尖波涟漪活动的低维表征与低维流形之间的关系——在距离上要比随机分布相隔低维流形要近,在时间上尖波涟漪的时间点要相邻的足够近。按照这样的标准,大部分的尖波涟漪发生在奖赏区域,约三成清醒阶段的尖波涟漪属于显著的重放。

图3. E 左:UMAP降维可视化与某次尖波涟漪事件降维位置(“位置流形”,按位置着色)。粉紫色点表示一个SPW-R重播事件的七个连续时间段的神经嵌入(每两个点间隔20毫秒);右:迷宫跑步期间的活动轨迹。黑色三角形表示重放事件发生时老鼠的物理位置。
利用之前介绍的解码方法,研究人员专门观察脉冲数据中的尖波涟漪活动。在原始高维空间中,该解码技术能够明确识别出尖波涟漪重放的是哪一个试次块。即使更换不同的距离度量和降维方法,解码结果也显示出了良好的一致性。在同一尖波涟漪事件中,不同时间点解码出的试次信息保持一致;但当减少参与解码的神经元数量时,这种一致性会迅速降低。这些结果均有效证明了这些解码方法在解读尖波涟漪期间神经元群体表征的时间信息方面是有效的。
最后,我们终于可以确定这些尖波涟漪重播的是过去、未来或是当前的试验块。通过用尖波涟漪本身发生的试次数减去解码出来它重放的试次数,便可以检查SPW-Rs的脉冲序列内容最可靠地解码到什么时间的试验块。结果清楚表明这个差值不偏不倚出现在零点,说明从尖波涟漪中解码出来的表征对于当前试次是最为可靠的。
在对研究方法进行深入探索,并在清醒状态下的尖波涟漪数据上验证之后,研究人员进一步探讨了清醒与睡眠阶段尖波涟漪之间的联系。通过将经历迷宫前的睡眠数据作为对照,我们发现,事后睡眠阶段的数据解码出的试次数分布与训练中尖波涟漪数据解码出的试次数分布高度一致。这一发现表明,在训练后的睡眠阶段,小鼠的神经元群体确实在重放训练期间所经历的事件。
但在训练过程中有那么多的因素影响,是什么因素使得事后睡眠脉冲序列解码出来的试次分布呈现这样的形态呢?换句话说,究竟是什么因素(包括迷宫中的重放、theta振荡强度、theta振荡波动次数、睡眠前的重放、随机噪声等等)使得某些试次会被在睡眠时重放呢?利用混合效应线性回归,研究人员发现在各种解码方法中,最显著的影响因素是迷宫中的尖波涟漪重放。
事件后睡眠与迷宫中的尖波涟漪除了解码出来试次数分布高度相关,还拥有同样的左右臂重放分布差异;这从空间解码的一致性上,验证了事后睡眠也许真的是在重放在迷宫中尖波涟漪发生时的神经活动。
未来展望:完整破解长期记忆形成的奥秘
在进行了以上一番探索性的工作之后,研究人员对最初的研究问题提出了答案。根据以往的研究,环境探索涉及到清醒活动与休息阶段之间的自然交替,这些变化需要大脑状态的相应调整,并且与尖波涟漪(SPW-Rs)期间的记忆重放紧密相关。
他们提出清醒状态下的尖波涟漪是一种自然的经验标记机制。这种机制通过对特定神经模式进行标记来实现,可能是通过将这些神经模式与先前的经验进行比较,并将其与动物的行为相关性连接起来。这些标记的模式在活动结束后的睡眠期间通过SPW-Rs多次重新激活,以此巩固选定的经验,并将其与大脑现有的知识基础结合。
此外,本文所发现的神经生理学机制还可能与之前的分子机制相关联。清醒状态下的尖波涟漪可能触发长期突触变化,从功能层面看,这可能形成一个依赖于大脑状态的神经元群体吸引子。当海马网络恢复到类似的大脑状态,如非快速眼动睡眠期间,由清醒状态下的尖波涟漪设定的模式便会再次被激活。
这种假说得到了以往研究的支持,研究表明,尖波涟漪能够影响那些与未来行为成功相关的神经活动,使其偏向于尖波涟漪时期的活动模式。例如,在迷宫中学习特定路径的老鼠,可能会记住路径上某些地点的特定外观、味道或声音作为成功获取奖励的线索。
在学习过程中,大脑可能会将这些线索与尖波涟漪事件关联起来,以帮助记忆的巩固并影响未来的行为选择。在人类中,非快速眼动(NREM)睡眠期间未被重新激活的记忆项目往往会被遗忘,而记忆的重放可以在单次学习后发生,且学习后的尖波涟漪数量可以预测醒来后的记忆表现。
总结而言,这篇文章表明清醒状态下的尖波涟漪是记忆选择的一种神经生理机制。这些发现为系统神经科学建立了一个涉及神经资源分配、表征漂移、事件重映射、时间编码和记忆编辑等多个领域的神经生理学框架。这一框架可能还可以与生物学和机器学习中的重要问题联系起来。例如,在机器学习中,重要性采样是一个关键技术,对其进行改进后,我们可能会实现更高的效率和更强的泛化能力。
参考文献
[1] FREY U, MORRIS R G. Synaptic tagging and long-term potentiation [J]. Nature, 1997, 385(6616): 533-6.
[2] JADHAV S P, KEMERE C, GERMAN P W, et al. Awake hippocampal sharp-wave ripples support spatial memory [J]. Science, 2012, 336(6087): 1454-8.
[3] FERNÁNDEZ-RUIZ A, OLIVA A, FERMINO DE OLIVEIRA E, et al. Long-duration hippocampal sharp wave ripples improve memory [J]. Science, 2019, 364(6445): 1082-6.
[4] STARESINA B P, ALINK A, KRIEGESKORTE N, et al. Awake reactivation predicts memory in humans [J]. Proc Natl Acad Sci U S A, 2013, 110(52): 21159-64.
[5] MACKEVICIUS E L, BAHLE A H, WILLIAMS A H, et al. Unsupervised discovery of temporal sequences in high-dimensional datasets, with applications to neuroscience [J]. Elife, 2019, 8.
[6] MCINNES L, HEALY J. UMAP: Uniform Manifold Approximation and Projection for Dimension Reduction [J]. ArXiv, 2018, abs/1802.03426.
本文来自微信公众号:追问nextquestion (ID:gh_2414d982daee),作者:赵诗彤,编辑:存源