
本文来自微信公众号:nextquestion(ID:gh_2414d982daee),作者:作者:物离,编辑:夏天,审稿:EY,原文标题:《研究大脑中的“GPS”,能帮我们破解神经元的算法吗?》,题图来自:《超体》
一、神经元的“算法”:频率编码还是时间编码?
在小说《三体》的设定中,外星文明“三体”世界构建出由三千万人组成的人列计算机,并尝试通过该计算机去预测其文明所依赖三颗恒星的运行轨迹。我们知道,对于计算机来说,算法是联接输入和输出的桥梁,只要选择好合适的信息输入,配上正确的算法便能输出特定的目标。即使是这个人列计算机,通过由两到三名士兵(每名士兵相当于一个二进制字符)协同工作形成的逻辑门,再配合上微分方程算法,也能够进行极大运算量的工作。
然而,面对数以百亿计的神经元组成的“人脑计算机”,我们通常能够直接观测到的只是输入刺激和输出反应的信息,对从刺激到反应之间发生了什么——也就是“神经元的算法”却知之甚少。神经科学家们只能根据刺激和反应的信息特征,来推断刺激和反应之间的映射关系,而这便是神经科学家们一直关心的最重要问题之一。
要破解“神经元的算法”,神经科学家们一般会采取两种手段,要么建立从刺激到反应的映射,着眼于理解神经元如何对不同的刺激做出相应的反应,进而建立模型来预测神经元对特定刺激的反应;要么研究相反方向的映射,从已知的神经元反应来推算外界刺激的面貌,试图从被激发的神经元活动序列来重构外界刺激或者刺激的某些特征。
那么面对不同刺激时,大脑中的神经元活动模式为什么能够编码信息?我们已经知道,神经元会因为刺激而产生动作电位,动作电位作为神经活动依靠突触完成细胞间的信息传递。而动作电位的产生有一个特点,那就是依赖于特定的阈限,表现出“全或无”的特征。这种“全或无”的特征就像计算机的二进制语言一样,要么是“0”,要么是“1”。在一段时间内,对神经元发放与否的记录,就能得到不同的神经元活动模式。
我们不妨再深入一点,想想对神经元处于0或1状态的特征检测,能记录到哪些信息?很容易想到的就是,一些神经元对某类刺激响应得多,对另一类刺激响应得少,那么神经元对某类刺激在一段时间内的发放次数不就是独立于特定刺激神经特征吗?
没错,神经科学家们也是这么干的,并把这种编码方式称作频率编码假说。他们认为信息是由神经元单位时间内产生的动作电位尖峰数量来编码的,并把这种指标称为神经元发放速率(Spike-count rate)。
但是频率编码也可能存在一定问题。举个例子,对于某个神经元来说,在一段时间内可能存在两种动作电位模式,有可能是1011011010,或者也有可能是1101101101(1代表有,0代表无)。根据频率编码假说,这两种神经元活动模式编码的信息相同,在这段时间内都产生了7次尖峰。但是如果神经元发放的顺序也包含着信息,那么用神经元发放速率却区分不出这两种信息的区别。所以神经科学家们也提出了时间编码假说,认为信息是由神经元尖峰产生的特定时间点来编码的,也就是看在接受刺激后某个特定的时刻,神经元是否发放。
其实频率编码假说和时间编码假说都得到了一些实验证据的支持,在这些实验中,“神经元的算法”依赖于特定的实验刺激和实验任务,并不统一。目前学界也没有定论,认为神经元对某类刺激就一定会采取某种特定的编码方式。因为真实的环境刺激复杂而又多变,所以神经科学家们想要破解神经元的“终极算法*”还道阻且长。不过近十年来,科学家们正在通过研究神经元对空间位置信息的编码,逐步揭开这个难题。
*注:“终极算法”的说法来源于美国作家佩德罗·多明戈斯所著的科普读物《终极算法:机器学习和人工智能如何重塑世界》,这里的“终极算法”是作者所构想的一种能够普遍解释智能对象学习机制的算法。
二、对空间位置的神经编码
知道自己正身处何方,清楚我们要去往何处,如何在复杂的环境中定位和导航是我们适应生存的必需技能,也是我们日常生活中最常被调用的认知能力。科学家们对于这一能力的研究,最早是基于大鼠或小鼠在迷宫中探索任务而展开的。
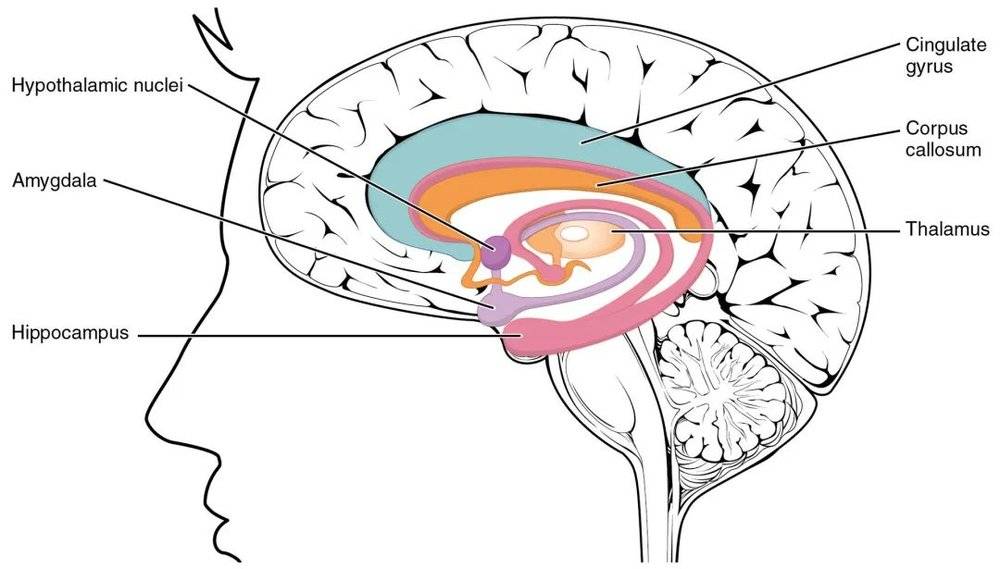
研究发现,鼠类脑中海马体和内嗅皮层中特定神经元的发放速率和它们在迷宫中的位置密切相关。也就是说,它们在迷宫中对空间位置的认知和导航是由特定的神经元活动编码的。2014年诺贝尔生理学或医学奖就颁发给了对该项工作做出杰出贡献的三位科学家,以表彰他们关于大脑中的“GPS”——位置细胞和网格细胞的相关成果。

当然,仅仅破解小鼠脑中“神经元的算法”,还不能让神经科学家们很好地回答我们最开始提出的问题,研究者在“人类的近亲”——灵长类动物身上重复了类似的发现。有了前面的这些基础之后,神经科学家们总要回归最终的问题,那就是人脑中的神经元对空间信息的表征是否也与之相似?
要想了解人类的某些特定的神经元如何对外界刺激进行反应,研究者们不能像对待实验动物那样,直接将记录电极插入人脑。
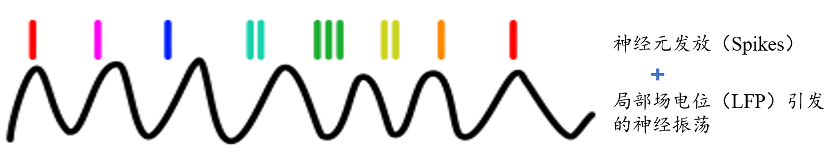
不过,一些志愿者在接受神经外科治疗时就已经在颅内安放了用于医学用途的电极,正是这些设备得以让神经科学家们观察到人类被试在进行相关任务时颅内的神经活动。被颅内脑电记录的神经活动,往往是通过一簇神经元的同步活动所引发的局部场电位(LFP,Local field potential)而产生的有节律的、周期性活动模式,即神经振荡(Neural oscillation)。而近两年的研究表明,人脑对空间导航和环境信息的编码,正是通过海马体、内嗅皮层或内侧颞叶的theta神经振荡*完成的。
*注:theta波是人类脑电波的重要成份之一,频率为3~8Hz,theta神经振荡是指频率为3~8Hz的周期性神经活动。
2020年发表在《自然-通讯》上的一项研究让被试在虚拟环境中运动的同时,记住沿路的物品信息。研究者调节了被试们在虚拟环境中的运动速度,这样被试就必须对他某个时刻所处的位置倍加留心。有趣的是,在虚拟运动过程中,被试的海马体后部出现了频率约为8Hz的神经振荡,而且振荡的精确频率和其运动速度密切相关。
也就是说,人类海马体中的部分神经元产生振荡活动的频率,是根据我们运动过程中位置的变化而变化的,这种对应关系就可以看作是一种“神经元的算法”。所以,研究者认为这个频段的theta振荡可能反映了被试的空间导航编码。与此同时,在被试的海马体前部也检测到了较低频的theta神经振荡,振荡的精确频率却和被试的运动速度无关,研究者认为其可能反映了海马体中的部分神经元对非空间信息的编码。
图片来源:OpenStax College
在日常生活中,我们不仅需要对自己的位置精确编码,知道我们在哪;有时候我们也需要精确地定位环境中其他个体的位置和环境的边界信息,知道我们要去哪。
2020年年末,《自然》上的一篇文章对该问题进行了探究。在这项研究中,被试需要在真实的物理环境中执行空间导航任务。这有点像小鼠玩的迷宫游戏,只是人类被试对自己要到达的目标位置更明确。结果发现,被试内侧颞叶出现的theta振荡同样能够编码他们所在的具体位置,同时还有环境中的目标位置和边界信息,并且后者和更高频的theta振荡有关。
而在今年6月发表在《细胞》的一篇文章中,Qasim等人报告了一种能够编码空间位置信息的神经元活动模式“相位进动”(Phase precession)的最新发现。相位进动不同于神经元发放速率和神经振荡,但却又和它们密切相关,它描述了大脑一个区域内大范围的神经振荡和单个神经元产生的尖峰电位之间的关系。
这就好像一个人绕着操场跑步的时候,总在操场边的长椅上看到一对小情侣在亲热,又总是在另一边的大树下看到一位小哥练习演讲,这里不管是情侣还是小哥的行为就好比特定神经元发放的尖峰,而跑步者的周期性运动就好比神经振荡。由于相位描述了周期波形中随着时间变化波的特性,所以这些特定的事件总是发生在一个周期内的特定时刻,对应着特定的相位(角),所以这样的一种神经元活动现象也就被称作相位进动。
图片来源:Qasim, S. E., Fried, I., & Jacobs, J. (2021). Phase precession in the human hippocampus and entorhinal cortex. Cell, 184(12), 3242–3255.e10.
这项研究结果发现,人类的海马体和内嗅皮层神经元的相位进动现象,也能够精确地编码被试的位置信息。并且研究者还进一步发现了与特定目标状态相关的相位进动现象,研究者甚至兴奋地指出:“这可能是个体空间导航,乃至表征学习和记忆中的顺序事件的一般神经模式”。
三、海马体中的相位进动
其实神经元活动中的相位进动现象,并非第一次被科学家所观察记录到。早在1993年,就有研究者描述了大鼠在运动过程中,海马区域的theta振荡和该区域神经发放率之间的关系。这篇发表在《海马》期刊的研究发现,海马神经元的发放总是开始于一个振荡周期中某个特定的相位,而不同的神经元发放开始的相位角也不尽相同。
在2009年的另一项发表在《神经科学期刊》的研究中,研究者则是直接检查了神经外科患者颅内神经振荡引发的局部场电位和神经元发放峰值之间的关系,从数学统计上找到了两者之间潜在规律。而在最近这项发表在《细胞》的研究中,研究者在特定的实验任务中,进一步细化了这种神经活动模式。
重要的是,这项最新的工作在人类的海马体和内嗅皮层观察到的相位进动,即被试处于特定的物理空间位置时,相应的神经元发放的峰值总在theta振荡的早期出现,和之前在啮齿类动物身上发现的实验结果非常相似,而这两个脑区正是一般被认为和我们日常的空间导航和情景记忆密切相关的脑区。
结合认知心理学相关的研究结果,一座架在微观、中观和宏观层面的桥梁得以显现,那就是海马体和内嗅皮层神经元的相位进动现象可能作为大脑对环境刺激信息的“算法”,让我们得以区分不同的空间位置信息,时刻清楚地了解自己身在何处,又要去往何方。
这项工作的发现无疑是令人激动的,除了结果本身的意义外,这项研究的价值还在于它为我们提出了一些新的问题。人类毕竟和小鼠不同,在这篇文章中研究者同时报告了一些和以往基于啮齿类动物实验的关键区别。其中最主要的便是在当前实验中,在人类身上发现的相位进动对空间位置信息的编码其实并没有小鼠那么明显,人类的theta波振荡频率也比小鼠要更慢、范围更广。研究者认为这可能和人类是在虚拟现实环境中,而非真实物理环境执行空间导航任务有关,并指出这可能正是之前没有在人类身上发现相位进动的原因之一。
笔者认为,这样实验结果也许蕴含了更多人类演化的意义。人类相比于其他物种的一个关键区别便在于,演化让人脑有了更大的可塑性,也就是其特定区域的功能并非是特定而专门的,而是可以泛化的。在该项研究中,研究者不仅发现了基于空间定位任务的相位进动,同时还发现了基于目标定向的相位进动,所以研究者才将相位进动的意义扩展至表征学习和记忆中的顺序事件的一般神经模式。
说到这里又不得不提到2020年一项发表在《认知科学进展》上的研究,研究者通过大量实验惊奇地发现,相比于其他物种,人类海马体中缺少模式分离*的机制,也就是说对于同一对象的不同形态,不管是图片、相似物、声音信息还是语言符号指代,都有相同的神经元对不同的刺激做出相同的反应模式。
笔者猜测,正是由于这种神经活动模式的“可塑性”,才导致了人类相比于其他物种拥有泛化能力、抽象能力以及创造能力这类进化上的优势。想象一下,如果一位尼安德特人面对一棵树的时候,只是惊叹于大树那历经岁月的参天轮廓,而想不到树叶可以用来遮体、树枝可以用来做武器、树干可以用来保存火源……那么也许下一秒他就会成为潜伏在树后面剑齿虎的腹中美餐。
*注:模式分离是指对不同的记忆对象,相应的神经元活动模式不同,是防止不同记忆相互干扰的神经编码的基本原则。模式分离的存在于不同物种间都得到了大量计算和实验证据的支持。
不过这种猜测在科学上当然是不负责任的,可能正如这项最新研究的作者指出的:“未来研究人类和其他物种之间相位进动的差异,对于我们进一步理解空间导航的神经编码机制至关重要”。假设,实验,比较,证实/证伪,毕竟这才是科学一贯的运作方式。有朝一日,也许我们真的能够完全理解我们大脑中神经元的工作方式,破解“神经元的终极算法”,相信那一定比灵动的计算机代码要更加精彩纷呈。
参考文献:
Qasim, S. E., Fried, I., & Jacobs, J. (2021). Phase precession in the human hippocampus and entorhinal cortex. Cell, 184(12), 3242–3255.e10. https://doi.org/10.1016/j.cell.2021.04.017
Stangl, M., Topalovic, U., Inman, C. S., Hiller, S., Villaroman, D., Aghajan, Z. M., Christov-Moore, L., Hasulak, N. R., Rao, V. R., Halpern, C. H., Eliashiv, D., Fried, I., & Suthana, N. (2021). Boundary-anchored neural mechanisms of location-encoding for self and others. Nature, 589(7842), 420–425. https://doi.org/10.1038/s41586-020-03073-y
Goyal, A., Miller, J., Qasim, S. E., Watrous, A. J., Zhang, H., Stein, J. M., Inman, C. S., Gross, R. E., Willie, J. T., Lega, B., Lin, J. J., Sharan, A., Wu, C., Sperling, M. R., Sheth, S. A., McKhann, G. M., Smith, E. H., Schevon, C., & Jacobs, J. (2020). Functionally distinct high and low theta oscillations in the human hippocampus. Nature communications, 11(1), 2469. https://doi.org/10.1038/s41467-020-15670-6
Quian Quiroga R. (2020). No Pattern Separation in the Human Hippocampus. Trends in cognitive sciences, 24(12), 994–1007. https://doi.org/10.1016/j.tics.2020.09.012
Buzsáki, G., Anastassiou, C. A., & Koch, C. (2012). The origin of extracellular fields and currents--EEG, ECoG, LFP and spikes. Nature reviews. Neuroscience, 13(6), 407–420. https://doi.org/10.1038/nrn3241
Manning, J. R., Jacobs, J., Fried, I., & Kahana, M. J. (2009). Broadband shifts in local field potential power spectra are correlated with single-neuron spiking in humans. The Journal of neuroscience: the official journal of the Society for Neuroscience, 29(43), 13613–13620. https://doi.org/10.1523/JNEUROSCI.2041-09.2009
O'Keefe, J., & Recce, M. L. (1993). Phase relationship between hippocampal place units and the EEG theta rhythm. Hippocampus, 3(3), 317–330. https://doi.org/10.1002/hipo.450030307
本文来自微信公众号:nextquestion(ID:gh_2414d982daee),作者:物离,编辑:夏天,审稿:EY