
本文来自微信公众号:Nature Portfolio(ID:nature-portfolio),作者:Stuart M. Brierley,头图来自:视觉中国
慢性腹痛很常见,但其背后的原因尚不清楚。新的研究发现,肠道感染会触发局部免疫反应,致使正常情况下无害的食物被当成“敌人”攻击,引起肠道持续疼痛。
疼痛机制的不断演化提醒并保护我们免受实际或可能的组织损伤。疼痛有三种常见形式:当机体检测到伤害性刺激时的伤害性疼痛,与炎症或感染相关的炎性疼痛,以及需要长期忍受的慢性疼痛[1]。Aguilera-Lizarraga等人[2]在《自然》上发表的研究基于来自小鼠和人的证据,报道了可能导致慢性肠道疼痛的一个新机制。
当一个人受到明显外伤,例如手臂骨折时,我们很容易就知道他们肯定很痛。但如果受伤部位不那么明显,疼痛的来源就很难确定了。这个问题在有内脏疼痛的人群中很常见,他们的疼痛有些还容易诊断,有些则容易成为疑难杂症。
举个例子,炎症性肠病的诊断指征可能很明显,如胃肠道出血、肠壁炎症以及粪便或血液样本中存在特定的生物标志物分子[3],而肠易激综合征(IBS)这种影响全球11%人口[4]的疾病却缺乏这些明确的特征,也没有明显的致病机制可以解释其患者经历的慢性腹痛和并发的便秘或腹泻。
既往研究表明,肠易激综合征在女性中更普遍[4],其症状可以由应激[4]、感染性肠胃炎(摄入不洁食物或水引起)[4]、肠道微生物组成改变[5],以及脑-肠轴功能异常[6]引起。现在,Aguilera-Lizarraga等人的研究表明,肠道细菌感染可以显著改变肠道中的局部免疫反应,这会让某些食物被当成是有害的,进而导致持续的肠道疼痛(图1)。
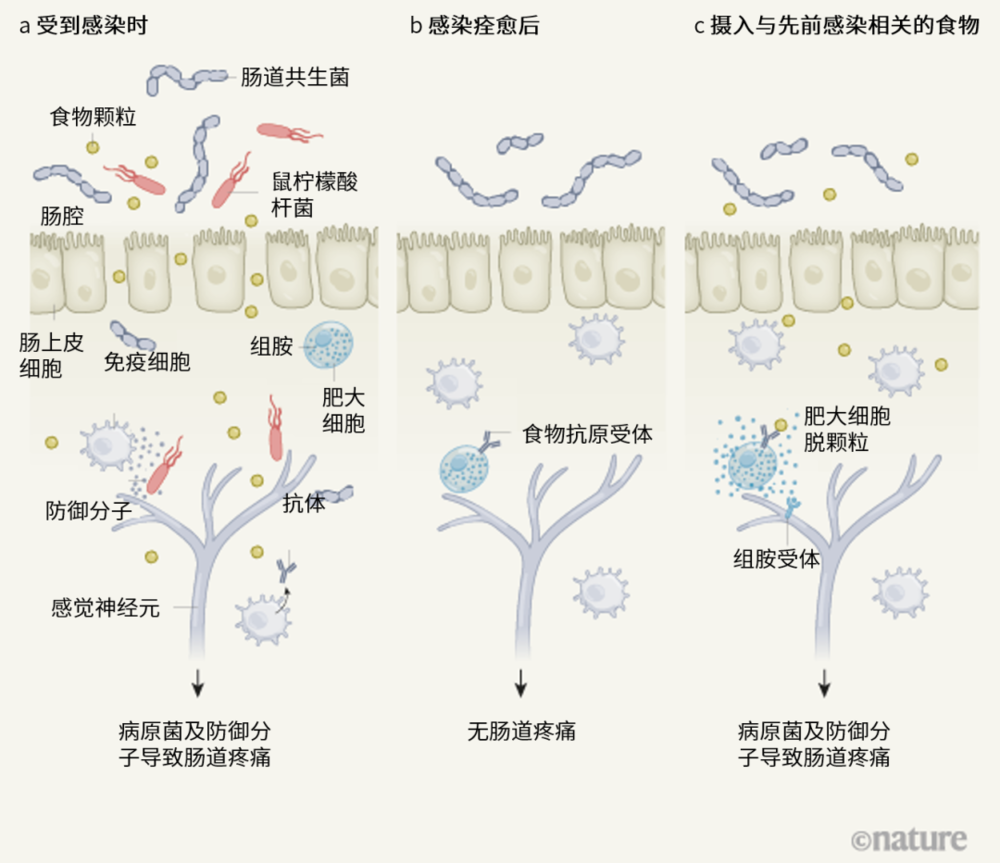 图1 | 无害食品引起的免疫反应可能导致疼痛。 Aguilera-Lizarraga等人[2]揭示了可能导致慢性肠道疼痛的新机制。a,包括含有组胺分子的肥大细胞在内的肠道中的免疫细胞通常不会针对食物或肠道共生菌产生免疫反应。但如果小鼠感染鼠柠檬酸杆菌,当肠黏膜屏障受损(肠道上皮细胞间连接被破坏)时,食物和细菌会离开肠道腔进入人体,而免疫细胞就会作出相应的反应。免疫细胞针对鼠柠檬酸杆菌释放防御分子,同时也可能针对无害的食物产生抗体。这会让感染导致肠道疼痛。b,当感染痊愈后,肠壁开始修复。肥大细胞会向神经元靠近,同时表达能够识别食物片段(即“抗原”)的受体(基于先前的抗体而产生)。c,此后如果再摄入这类食物,肥大细胞会作出相应识别,并通过脱颗粒的方式释放组胺。组胺与相应受体结合,激活感觉神经元,进而引起疼痛。
图1 | 无害食品引起的免疫反应可能导致疼痛。 Aguilera-Lizarraga等人[2]揭示了可能导致慢性肠道疼痛的新机制。a,包括含有组胺分子的肥大细胞在内的肠道中的免疫细胞通常不会针对食物或肠道共生菌产生免疫反应。但如果小鼠感染鼠柠檬酸杆菌,当肠黏膜屏障受损(肠道上皮细胞间连接被破坏)时,食物和细菌会离开肠道腔进入人体,而免疫细胞就会作出相应的反应。免疫细胞针对鼠柠檬酸杆菌释放防御分子,同时也可能针对无害的食物产生抗体。这会让感染导致肠道疼痛。b,当感染痊愈后,肠壁开始修复。肥大细胞会向神经元靠近,同时表达能够识别食物片段(即“抗原”)的受体(基于先前的抗体而产生)。c,此后如果再摄入这类食物,肥大细胞会作出相应识别,并通过脱颗粒的方式释放组胺。组胺与相应受体结合,激活感觉神经元,进而引起疼痛。
健康人中,口服耐受(oral tolerance)会让免疫系统“忽略”从口进入肠道的物质[7,8]。这种耐受不包括被我们机体认定为有害的物质,如致病性细菌、寄生虫和病毒。我们的身体通过名为抗原的分子片段发现外来入侵者,该片段相当于一种“条形码”,能使我们的免疫系统特异性地识别入侵者。
我们的免疫系统会产生抗体,找到它们,从而对这些抗原加以标记,这样一来,一旦病原体再次出现,它们就会迅速成为攻击对象。我们体内的防御系统一般只攻击“坏人”,不会误伤无辜的“路人”。然而,根据Aguilera-Lizarraga和同事提出的假设,口服耐受失效会导致免疫系统不分敌我,胡乱攻击。
为了确定这种假设的免疫耐受失效是如何在小鼠体内发生的,研究团队利用鼠柠檬酸杆菌(病原菌)和卵清蛋白(蛋白中的一种蛋白质)构建了一个模型体系。研究人员评估小鼠肠道疼痛的方法,是测定小鼠结直肠扩张引起的腹部挛缩。多次摄取卵清蛋白并不会引起胃肠道疼痛;但小鼠从鼠柠檬酸杆菌感染中恢复后再次进食卵清蛋白,则会发生肠道疼痛和腹泻。
这些小鼠还会出现“肠道渗漏”的情况,即肠粘膜上皮通透性增加。换句话说,肠粘膜上皮细胞并未发挥其物理屏障的正常作用,导致肠道内容物进入粘膜下组织,进而引起了免疫反应并激活了感觉神经。此外,研究人员还在小鼠直肠内发现了针对卵清蛋白的特异性抗体,但其他部位并没有类似发现。
预防感染后小鼠肠道疼痛有两种方法:一种是对其进行基因改造使其缺乏IgE(一种抗体),另一种是给予小鼠抗IgE抗体,阻断其免疫系统产生的卵清蛋白特异性IgE抗体。相反,如果向未感染鼠柠檬酸杆菌小鼠的直肠注入卵清蛋白特异性IgE抗体,进食卵清蛋白后它们会出现与感染小鼠类似的肠道疼痛。
随后,Aguilera-Lizarraga和同事进一步阐明了这种疼痛反应背后的分子机制。他们的研究表明,在鼠柠檬酸杆菌感染后注射卵清蛋白,小鼠结肠中的肥大细胞(一种免疫细胞)会发生脱颗粒,这个过程会释放包括组胺在内的防御分子。通过药物阻断这一过程,无论是防止肥大细胞脱颗粒还是通过基因工程让小鼠彻底缺失肥大细胞,都可以显著减轻或是预防小鼠感染后进食卵清蛋白诱发的肠道疼痛。
研究人员还用证据表明,组胺的释放会影响肠道的感觉神经元,进而触发疼痛。从这些小鼠结肠提取的上清液在体外试验中能够提高疼痛感觉神经元的敏感性。通过药物阻断组胺H1受体或通过基因工程使小鼠缺乏这种受体,都可预防上述现象。
接下来,研究人员将与食物过敏有关的大豆、小麦、麸质、牛奶的混合液直接注射到12名肠易激综合征患者和8名健康个体的结肠直肠中;这些食物可以导致包括腹痛腹胀在内的肠道症状[7]。所有肠易激综合征患者表现出至少对其中一种食物的免疫反应迹象——而健康组中只有2人表现出这种迹象。与健康人群相比,肠易激综合征患者在神经纤维附近的肥大细胞更多,这表明肥大细胞与感觉神经元神经末梢之间的信息传递更为频繁有效。
研究人员报告称,23%的肠易激综合征患者粪便样本对金黄色葡萄球菌感染呈阳性,而健康受试者的粪便样本阳性率仅为9%。这一发现很有趣,因为金黄色葡萄球菌是“超级抗原”的主要微生物来源之一——所谓超级抗原是指与T细胞非特异性激活有关的抗原[9]。
事实上,47%的肠易激综合征患者的粪便样本中至少对一种超级抗原呈阳性,而健康受试者的粪便样本的阳性率仅为17%。这些发现表明,既往感染以及超级抗原的存在,可能通过诱发免疫反应加剧部分肠易激综合征患者的肠道疼痛。
本研究中有几个问题需要进一步考量深思。譬如,研究人员在小鼠中进行机制探讨时是让小鼠口服潜在抗原,而在人体试验中则是通过肠粘膜给予抗原刺激。口服上述可能致敏的食物是否会出现相同的研究结果有待进一步验证。
此外,这种“口服耐受失效”理论也无法解释为何肠易激综合征患者中女性比例显著高于男性[4]。尽管Aguilera-Lizarraga团队的研究似乎能揭示感染后发生肠易激综合征(譬如胃肠炎)的相关机制,但值得进一步探讨这种机制是否适用于其他类型的肠易激综合征,如便秘型、腹泻型、或腹泻-便秘交替型肠易激综合征。
本研究使用了鼠柠檬酸杆菌作为其小鼠模型中的致病菌。但其他有害微生物感染,如大肠杆菌、沙门氏菌、贾第鞭毛虫和志贺氏菌,都可能导致肠易激综合征。临床流行病学研究表明,肠道发生以上病原体感染都可能增加个体罹患肠易激综合征[10]的可能性。
这种机制是否仅适用于结直肠,还是在胃肠道其他区域(如胃,小肠和近端结肠)也存在?如果其他区域确实存在类似机制,那可能会激活不同的感觉神经,从而引发其他与肠道疼痛疾病相关的症状,例如恶心、腹部不适、腹胀等,功能性消化不良就属于这类疾病[4]。
本研究提出了诸多可供参考的治疗干预方案,包括改善肠屏障功能,减少肠道内容接触肠道免疫系统的机会;阻断相关食物特异性IgE抗体;减少肥大细胞脱粒;靶向肥大细胞释放的分子或其作用的受体;阻断传递有害信号并引起疼痛的结肠感觉神经。
口服耐受一旦失效,还能通过改变饮食习惯恢复吗?对此,食物过敏研究表明,从饮食中去除可能致敏的食物,再慢慢重新引入,可以改善长期预后[11]。如今,限制性饮食已经越来越多地用于治疗胃肠道症状,如无麸质饮食治疗乳糜泻;对于肠易激综合征,FODMAPs饮食限制了特定碳水化合物的摄入,这类碳水化合物在肠道中无法被完全消化吸收,FODMAPs指的是可发酵的低聚糖、二糖、单糖和多元醇[12]。
综上,Aguilera-Lizarraga团队的研究提出了与腹痛相关的新机制,为俗话说的“人如其食”再添有力证据。
参考文献:
1. Basbaum, A., Bautista, D. M., Scherrer, G. & Julius, D. Cell 139, 267–284 (2009).
2. Aguilera-Lizarraga, J. et al. Nature 590, 151–156 (2021).
3. Kobayashi, T. et al. Nature Rev. Dis. Primers 6, 74 (2020).
4. Enck, P. et al. Nature Rev. Dis. Primers 2, 16014 (2016).
5. Mars, R. A. T. et al. Cell 182, 1460–1473 (2020).
6. Grundy, L., Erickson, A. & Brierley, S. M. Annu. Rev. Physiol. 81, 261–284 (2019).
7. Nowak‑Wegrzyn, A., Szajewska, H. & Lack, G. Nature Rev. Gastroenterol. Hepatol. 14, 241–257 (2017).
8. Mowat, A. M. Nature Rev. Immunology. 18, 405–415 (2018).
9. Thammavongsa, V., Kim, H. K., Missiakas, D. & Schneewind, O. Nature Rev. Microbiol. 13, 529–543 (2015).
10. Spiller, R. & Garsed, K. Gastroenterology 136, 1979–1988 (2009).
11. Yu, W., Hussey Freeland, D. M. & Nadeau, K. C. Nature Rev. Immunol. 16, 751–765 (2016).
12. Moayyedi, P., Simrén, M. & Bercik, P. Nature Rev. Gastroenterol. Hepatol. 17, 406–413 (2020).
原文以Food for thought about the immune drivers of gut pain标题发表在 2021年1月13日的《自然》的新闻与观点版块上
本文来自微信公众号:Nature Portfolio(ID:nature-portfolio),作者:Stuart M. Brierley